Poster
# 108
![]()
Poster |
|
6th Internet World Congress for Biomedical Sciences |
Johan Van Doornik(1), Dimitris Patikas(2), Daniel Olivier(3), Gabriella Cerri(4), Michel Ladouceur(5), Jens Bo Nielsen(6), Thomas Sinkjaer(7)
(1)Centre for Sensory-Motor Interaction - Aalborg. Denmark
(2)Aristotle University of Thessaloniki - Thessaloniki. Greece
(5)Dept. of Medical Informatics and Image Analysis. Center for Sensory-Motor Interaction (SMI). Institute of Electronic Systems, Aalborg University - Aalborg O. Denmark
(7)Center for Sensory Motor Interaction. Aalborg University - Aalborg. Denmark
|
|
[Neuroscience] |
[Physiology] |
[Neurology] |
[Physical Therapeutics & Rehabilitation] |
|
The stretch reflex is one of the tools used in assessing neurophysiological changes over time. After a stretch of the ankle dorsiflexors, three distinct peaks (M1, M2, and M3) can be recognised in the EMG recorded from the Tibialis Anterior (TA) muscle. The first peak M1 reflects most probably the monosynaptic spinal reflex originating in the Ia afferents (Toft et al. 1989). The medium-latency response (M2) is mediated by afferent fibers with a slower conduction velocity; most likely spindle group II afferents (Schiepatti et al. 1997). The long-latency response (M3) is supposed to be - at least partly - mediated by a transcortical reflex. One of the evidences for this statement is shown by the facilitation that takes place in the PSTH of single TA motor units when the peaks evoked by transcranial magnetic stimulation (TMS) come at the same time as the M3 response (Petersen et al. 1998). TMS is a tool to study motor output in human subjects. The Motor Evoked Potentials (MEPs) are influenced by the excitablility of the motor cortex and the -motoneuronal pool (Rothwell 1997; Taylor et al. 1996).
In this study both changes in the size of each of the components of the stretch reflex and changes in the MEPs in participants performing a specific type of physical exercise (2 x 15 minutes balance training) are examined with the aim of finding a relation between this type of exercise and the occurring changes in the transmission that underlies the different pathways. Some evidence of a relationship between physical activity and the size of the monosynaptic H-reflex and the M1 component of the stretch reflex has already been presented (Casabona et al. 1990; Nielsen et al. 1993; Trimble and Koceja 1994), whereas it was also found that the M3 component was specifically increased in weightlifters (Mortimer and Webster (1983). Plastic changes in the motor systems of the spinal cord and cortex are also well documented from animal experiments (Wolpaw 1997; Nudo et al. 1997). Changes in the effect of TMS have also been demonstrated in relation to motor learning in human subjects (Cohen et al. 1998). For this study it was hypothesised that the required continuing attention of the participant to his or her balance would be reflected by a differential change in the size of the TA M3 component directly after the training session. In addition, it is expected that a differential change in M3 will be concomitant with a similar change in MEP.
Stretch reflex
The participants were seated in an adjustable chair. The right foot was strapped to a foot plate attached to a DC motor. A DC amplifier (Bruel and Kjær, Model 2708) powered the DC motor. The axis of rotation of the ankle joint and the foot plate were the same. The rotational velocity was controlled by a personal computer. The ankle torque was measured using strain gauges mounted on the connecting beam between the foot plate and the motor. The ankle angle was measured with a potentiometer.
A preliminary experiment consisting of establishing the optimal rotational velocity to measure the M3 component was effectuated. The effect of the stretch velocity on the amplitude of the stretch reflex components was investigated on four different participants. The subjects were asked to maintain a constant level of precontraction in the ankle dorsiflexors (5-10 Nm). The stretch reflex was recorded with 6 different velocities (12, 16, 25, 50, 85, 140 deg/s) and averaged over 40 times (after filtering as described above). The processed TA EMGs were then manually examined for the M1, M2 and M3 peaks. The amplitude was measured in relation to the background EMG level (50 ms average before stretch onset). In the balance training experiments the rotational velocity was set to 50 deg/s.
Transcranial magnetic stimulation
The transcranial magnetic stimulation was done with a Dantec Lite R-25 stimulator. The stimulus strength was manually adjustable on a scale from 0 to 100. A circular coil (Dantec NMC 140; Æ 70 mm) was used in order to elicit MEPs in both the TA and Soleus muscle. The peak-to-peak value of the short latency MEP as a function of the stimulus intensity was established for stimuli intensity ranging from 20% to 100% of maximal stimulator output. The data points were fitted with a Boltzmann sigmoidal curve. This was done by an iterative process, which minimised the sum of the squared errors (Prism, GraphPad Software Inc.).
Data acquisition
The TA and SOL electromyograms (EMG) were recorded with bipolar surface electrodes at a sample frequency of 2 kHz, after they were band pass filtered between 20 Hz and 1 kHz. The raw data was then rectified and low pass filtered with a 2nd order Butterworth filter at 20 Hz. The ankle torque and ankle angle were recorded with a sample frequency of 1 kHz.
Protocol
The effect of balance training on the stretch reflex and MEP was investigated in six different participants. They where asked to perform two sessions of 15 minutes balance training on a balance board used for rehabilitation, consisting of a bottom half sphere under a circular wooden plate. During the sessions the participants were encouraged to keep standing, but no clues where given regarding the technique. The number of errors where counted to monitor the learning process. The TA stretch reflex and MEP input-output curve were recorded before and immediately after balance training.
Preliminary stretch reflex experiments
Figure 1 shows the TA EMG, SOL EMG, ankle torque, and ankle angle at three different stretch velocities from participant 1. The TA EMG shows the M1, M2 and M3 peak. The SOL EMG remains very small during the stretch. The peak values M1, M2 and M3 from this and three other participants are measured at six different stretch velocities and are shown in figure 2. These results indicate that the M3 component is modulated by the stretch velocity and appears to be maximal for 50 deg/s. This stretch velocity will be used for the following experiments.
Balance training experiments
In figure 3 the TA EMG, SOL EMG, Ankle torque, and ankle angle during the TA stretch reflex are shown from before and after the balance training of participant 5. Surprisingly, the M3 component shows a decrease, whereas the M1 component remains the same. Figure 4 shows the MEP in the SOL and TA at 35%, 55%, 75% and 95% stimulus intensity before and after the balance training in the same participant. Figure 5 shows the peak-to-peak value of the MEP as a function of the stimulus intensity (the input-output curve). The reduction of the plateau phase of the fitted Boltzmann sigmoidal curve is significant within a 95% confidence interval.
Table 1 shows the results for 6 participants. Indicated is whether the stretch reflex components increased, decreased or remained the same, and whether the plateau phase of the MEP increased or decreased within a 95% confidence interval
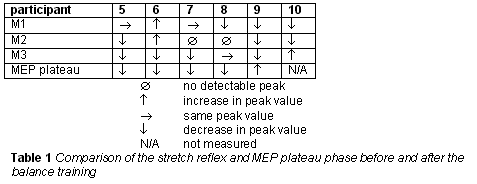
As presented in the title, these results are obtained during a pilot study. The results show only a limited number of subjects and hence should be taken cautiously.
Like other studies we found that physical exercise could modify the components of the stretch reflex and the size of the MEP (Casabona et al. 1990; Nielsen et al. 1993; Trimble and Koceja 1994; Mortimer and Webster 1983). However, our study is to our knowledge the first one that can reject the confounding factor of genetic predisposition.
At this point it is not clear what the influence of fatigue (Gandevia 1998) is on the differential changes in between the components of the stretch reflex results. This should be investigated further.
A change in the M3 component of the stretch reflex can be caused by several reasons, such as a change in gamma drive, presynatic or postsynaptic inhibition. Therefore we compared changes in the different components of the stretch reflex (M1 versus M3). A differential modification of the M1 versus M3 component indicates, most probably, that changes are originating from different neural structures than the ones involved in the monosynaptic reflex arc. Only two participants showed both a decrease in M3 and in the plateau phase of the MEP. This might be due to the complex relation between those two very different techniques in evaluating the cortical drive. However, the results indicate that one adaptation strategy for the balance board could be a decreased motor cortical influence, maybe to prevent overcompensation while balancing.
|
|
[Neuroscience] |
[Physiology] |
[Neurology] |
[Physical Therapeutics & Rehabilitation] |
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}