|
ESTRUCTURA HISTOLÓGICA DE LA CORTEZA CEREBELOSA
Prof. Dr. D. Héctor Fernández García
Área de Histología, Biología celular y Anatomía Patológica.
Dep. de Ciencias de la Salud. Universidad Rey Juan Carlos. Madrid
(España)
INTRODUCCIÓN
El cerebelo es un
órgano impar y medio, perteneciente al SNC, descubierto por Herófilo
en el siglo IV a.c. y alojado en la cavidad craneana, más
concretamente en la fosa posterior. A la simple inspección
macroscópica lo podemos dividir en vermis y hemisferios. Utilizando
la sistematización de Larsell (17) podemos describir el lóbulo floculonodular o arqueocerebelo, que está relacionado con el
vestibular y tiene como núcleo al núcleo del techo, Lóbulo anterior
o paleocerebelo, que recibe a los espinocerebelosos, gobierna el
tono postural y tiene como núcleo al globoso y emboliforme; lóbulo
posterior o neocerebelo que gobierna la motilidad voluntaria, recibe
aferencias del cerebro y tiene como núcleo al dentado. Por lo tanto,
el cerebelo es un órgano que se ocupa del ajuste y coordinación de
la musculatura estriada, tanto en sus funciones automáticas
(postura, etc.) como voluntarias, se puede tocar el piano gracias al
cerebelo. El patrón histológico de la corteza, es básicamente igual
en todas las áreas, y se puede considerar que es la suma de multitud
de unidades histofuncionales conocidas como laminillas.
CONCEPTO DE LAMINILLA CEREBELOSA
Si se efectúa un corte
sagital del cerebelo, se observa que la sustancia blanca se arboriza
(es el "árbol de la vida" de Herófilo). Las últimas arborizaciones
están cubiertas de sustancia gris formando las circunvoluciones y
surcos que se ven en la superficie externa, si observamos al
microscopio estas estructuras, notamos que cada tallo de sustancia
blanca se arboriza aún más, de forma que cada circunvolución está
integrada por microcircunvoluciones invisibles al ojo (fig.1). Estas microcircunvoluciones son las laminillas cerebelosas que se pueden
definir como formadas por un eje de sustancia blanca (que en
realidad es una lámina) recubierto por una banda de sustancia gris.
Estas laminillas, al igual que las circunvoluciones presentan dos
ejes, y ante un corte sagital del cerebelo, al más largo, que se
dispone perpendicular al plano del corte, se le denomina eje
longitudinal y al más corto, que se sitúa dentro del plano del
corte, transversal.
Arriba
ESTRUCTURA DE LA
LAMINILLA CEREBELOSA
Nos encontramos ante una estructura que presenta
un eje de sustancia blanca central rodeado por la sustancia gris
periférica que se denomina corteza y que acaba en la superficie del
órgano revestido por las meninges. En la corteza podemos diferenciar
dos capas de morfología muy diferente, que desde la sustancia blanca
hasta las meninges son la capa granulosa, granular o de los
granos y la capa molecular o plexiforme. En la inmensa
mayoría de los tratados se describen tres capas en la corteza
cerebelosa: granulosa, de las células de Purkinje y molecular,
aunque nosotros solamente consideramos la existencia de la primera y
la última, puesto que los somas de las células de Purkinje, como
veremos más adelante no forman capa continua y la célula está
ocupando todo el espesor de la corteza.
Efectivamente observamos, a partir de la
sustancia blanca una capa, la granulosa, que nos llama la atención
por estar ocupada por innumerables somas redondos de células
pequeñas, cuyos núcleos presentan cromatina relativamente densa, lo
que les confiere un aspecto linfocitoide. Por encima de la anterior
y hasta la superficie del órgano, se sitúa una banda con pocos
somas, la capa plexiforme, que presenta caracteres tintoriales
eosinófilos (en cortes teñidos con H-E) y que posee pocas neuronas
que se disponen salpicados en el espesor de la capa.
Con fines expositivos y aunque luego
describiremos cada tipo celular en detalle, las variedades
neuronales de cada capa son las siguientes:
CAPA GRANULAR: -Granos.
-Células de Golgi.
-Células de Lugaro.
-Célula monodendrítica o
monopolar en penacho.
CAPA MOLECULAR: -Células
estrelladas.
-Células en cesta.
En el límite entre las dos capas, llama la
atención la disposición de los somas de unas neuronas grandes, las
células de Purkinje, que se sitúan alineados y algo separados
entre sí y que ocasionaron que la mayoría de los tratados lo
consideren una capa individualizada. Estas células son posiblemente
las más importantes de la corteza y sus axones, como luego veremos
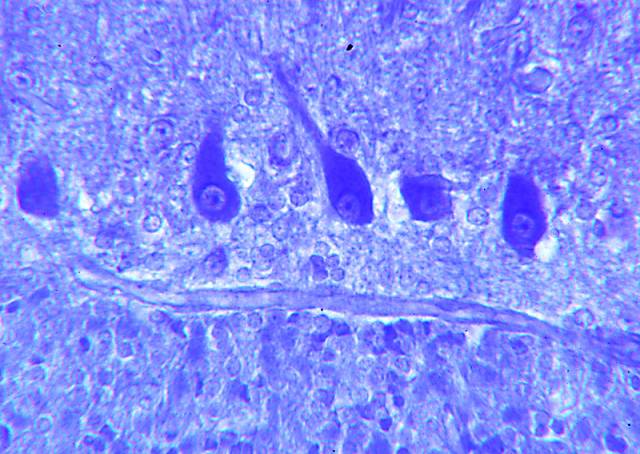
constituyen la única vía eferente de la misma (fig. 3).
Arriba
CÉLULA DE PURKINJE.
SOMA:
Las células de Purkinje,
fueron descritas en 1837 en Praga por Juan Evangelista
Purkinje, gracias a que fue el primero que pudo lograr cortes
histológicos de los centros nerviosos. Estas células, que son
más abundantes en la convexidad de la laminilla que en la
profundidad del surco, sitúan sus somas con una disposición
geométrica en rombos de aproximadamente 50 µm. de lado. Tienen un
soma grande (40-100 µm.), piriforme, algo aplastado en el eje
longitudinal de la laminilla y presentan un gran núcleo central,
esférico y de cromatina laxa en el que se muestra un nucleolo muy
patente. Su citoplasma es rico en grumos de Nissl que a veces se
disponen en paquetes perinucleares, neurofibrillas que forman una
red que penetra en el espesor de las dendritas, un diplosoma y un
complejo de Golgi bien desarrollado. (figs. 3, 4
y 5).
La M.E. confirma en lo básico esta descripción.
El núcleo es esférico, a veces escotado y cromatina difusa con un
nucleolo prominente. En el citoplasma se observan mitocondrias, un
complejo de Golgi muy desarrollado y gran cantidad de R.E.R.
dispuesto en paquetes, ya que son células con un gran árbol
dendrítico y mucho recambio protéico y de membrana. También
observamos en su citoplasma lisosomas que aumentan con la edad,
neurotúbulos y neurofilamentos no tan abundantes como sugiere la M.O.,
abundante R.E.L. y cisternas hipolemnales que si bien no son
específicas de esta célula, pues son habituales en todas las
neuronas grandes, son tan abundantes que se podrían considerar un
dato patonogmónico y que también se encuentran en los grandes
troncos dendríticos. Con mucha frecuencia se observa una mitocondria
en relación con estas cisternas hipolemnales y su significación
funcional no está clara pero se ha asociado con el recambio de
membranas y con la captación de sustancias externas hacia el
citoplasma neuronal. (figs. 6, 7
y 8).
Recientemente se han efectuado estudios
inmunohistoquímicos de las neuronas de la corteza (Hawkes y Leclerc,
1989 (10); Sahin y Hockfield, 1990
(34);Yan y Garey, 1996
(39),
etc.) y se demuestra que las células de Purkinje son positivas a los
anticuerpos anti-calbindina, y mabQ113.
DENDRITAS:
El árbol dendrítico se encuentra en su totalidad
incluido en la capa molecular, Nace de uno o dos gruesos troncos
principales que arrancan de la zona superior del soma y se arborizan
en el plano transversal de la laminilla, de forma que su
ramificación solo se observa en cortes transversales, mientras que
en los longitudinales no se aprecia más que una línea ascendente que
casi alcanza la superficie del órgano. Por lo tanto, el campo
anatómico que cubre el ramaje dendrítico es muy amplio en el plano
parasagital pero muy estrecho en el longitudinal. En otras palabras,
las células de Purkinje dejan abundantes áreas sin cubrir.
(figs.
3, 5, 9
y 15).
Estas dendritas se dividen en ramas cuyo diámetro
va disminuyendo progresivamente y forman las ramas dendríticas
secundarias y terciarias. De estas últimas surgen, a su vez, lo que
Cajal denominó "ramos terminales", que son más cortos, se dividen
solo una o dos veces y son de calibre constante. Sin embargo el dato
más llamativo de estos ramos terminales es que su superficie está
repleta de espinas sinápticas lo que les confiere un aspecto como de
"alambre de espinos".
Las dendritas principales contienen gran cantidad
de neurotúbulos, todos dispuestos de forma paralela y longitudinal y
condicionando la disposición de las restantes estructuras, de las
cuales las mitocondrias, que se suelen disponer periféricamente, son
lineales, el complejo de Golgi suele ser axial y longitudinal, así
como el R.E.L. que es muy abundante. El R.E.R. es escaso como ya
demostró Cajal y son abundantes las cisternas hipolemnales en las
que es frecuente su disposición en pilas de monedas.
En las espinas, por el contrario no se encuentran
mitocondrias ni neurotúbulos y solo contienen algunas cisternas de
R.E.L.. Estas espinas se han estudiado exhaustivamente porque, como
ya postuló Cajal, representan estructuras postsinápticas.
Habitualmente son formaciones "en tachuela", con un pedículo de
1'5-2 y una cabeza de 0'5 µm. de diámetro donde recibe la aferencia.
Puesto que esta estructura es perfectamente demostrable con el
método de Golgi, se ha intentado hacer análisis cuantitativos para
conocer el número de aferencias que a traves de las espinas recibe
la célula de Purkinje. El resultado de estos contajes varían, ya que
con el método de golgi solo se observan las espinas que se sitúan de
perfil, es decir, aproximadamente la mitad o un tercio del total de
las existentes. Considerando este factor de corrección, se calcula
que cada célula de Purkinje en la rata posee de 30.000 a 60.000
espinas, lo que es coincidente con los contajes de Fox
(8) en el
mono y de Palkovits (28) en el gato y discrepante con resultados de
otros autores, aunque en cualquier caso sirven para dar una idea de
la tremenda cantidad de aferencias que reciben las células de Purkinje. En los troncos dendríticos principales y secundarios,
existen unos abultamientos sin tallo, mayores y más groseros que las
espinas, y que también serán localizaciones para recibir contactos
sinápticos.
AXÓN:
El axón de las
células de Purkinje es un elemento de gran importancia para la
comprensión de la corteza cerebelosa, ya que es la única vía por la
que se canalizan las eferencias. El axón surge del soma, en
situación contrapuesta a las dendritas, o bien más raramente de uno
de los troncos dendríticos principales, dirigiéndose mediante un
trayecto ligeramente arqueado a la sustancia blanca. A diferencia de
otras neuronas no existe un cono axónico definido, de manera que,
como ya dijo Cajal (4), el único dato diferencial es la convergencia
de las neurofibrillas. Su segmento inicial mide 1-2 µm. de diámetro
por 15-20 de longitud, de forma que se extiende por la parte
superior de la capa de los granos, es amielínico y no da
colaterales. Su límite está señalado por un adelgazamiento brusco,
durante el cual se hace casi imperceptible (0'2-0'4 µm. de
diámetro), volviendo luego a engrosarse y mielinizarse; una vez que
ha adquirido la envoltura mielínica atraviesa la capa de los granos
e ingresa en la sustancia blanca, terminando en los núcleos grises
del cerebelo. Antes de abandonar la capa de los granos y una vez
mielinizado, da una colateral recurrente descrita por Cajal, a
partir de la cual van a formarse varios importantes sistemas de
asociación.
Esta colateral, cerca de su origen, da una
rama que forma un plexo relativamente poco extenso en la capa
granulosa, como ya describiera Lugaro y posteriormente comprobaron
Fox (8) y Palay (27). Una vez que ha surgido esta rama, la colateral
sigue un trayecto ascendente recubierta por su vaina de mielina y se
resuelve a la altura de los somas de las células de Purkinje en dos
plexos, uno situado por debajo (infragangliónico) y otro por encima
(supragangliónico) que terminan sobre distintos somas neuronales,
entre los que se encuentran los de otras células de Purkinje. En
cuanto a la porción axónica que ingresa en la sustancia blanca,
termina en un núcleo cerebeloso formando un teledendrón en tirabuzón
que abraza varias de las grandes neuronas ganglionares. El análisis
E.M. de estas terminaciones ha revelado que se trata de contactos
axo-somáticos y axo-dendríticos muy extensos y al igual que los de
las colaterales, de tipo II de Gray y con vesículas planas, lo que
está en correspondencia con su carácter inhibidor. Sin embargo, este
carácter inhibitorio es sorprendente, pues si resulta que la única
aferencia de la corteza cerebelosa tiende a inhibir, ha de admitirse
que las neuronas nucleares reciben influencias activadoras de otro
origen.
Arriba
CAPA DE LOS GRANOS.
Como ya comentábamos más arriba, esta capa nos
llama la atención por estar ocupada por innumerables somas redondos
de células pequeñas (granos del cerebelo), cuyos núcleos presentan
cromatina relativamente densa, lo que les confiere un aspecto
linfocitoide. y en la que de trecho en trecho llaman la atención
unos pequeños espacios acelulares eosinófilos a los que Cajal
denominó "Islotes protoplásmicos" (fig. 14) y en los que Held
demostró "neurosomas" (mitocondrias), los cuales tendrán una
significación funcional muy importante. Esta capa se extiende hasta
aproximadamente la mitad de la corteza, teniendo unos 500 µm. de
espesor en la convexidad y unos 100 µm. en el surco. Como ya hemos
apuntado más arriba es la más celular y se calcula que contiene de 3
a 7 millones de neuronas por mm3. El elemento celular
predominante es el grano del cerebelo. Junto a ellas existen otros
elementos más grandes conocidos como células estrelladas grandes y
que constituyen un tipo celular alrededor del cual existe un notable
confusionismo terminológico. (figs. 1
y 3).
GRANOS.
Son abundantísimos y el elemento más
característico de esta capa, calculándose que la relación células de
Purkinje/granos es de 1/1769 (29) en el gato. Su soma es esférico y
pequeño (entre 5 y 8 µm. de diámetro), no presentan grumos de Nissl
y su núcleo, ocupa la mayor parte del citoplasma y presenta
cromatina densa, lo que provoca una gran cromofilia. Estas dos
características prestan a la célula un aspecto linfocitoide. De
hecho en 1900 todavía se dudaba que no fueran linfocitos. Los somas
están muy juntos unos a otros y pese a que no están revestidos por
glía no presentan contactos sinápticos. El núcleo es ligeramente
irregular, con heterocromatina en bloques y en el citoplasma se
observan algunas mitocondrias, un pequeño complejo de Golgi,
polisomas y un diplosoma ya descrito por del Río Hortega.
(fig. 10,
11 y 12).
Las dendritas son 4 o 6 que nacen del soma y
tras un trayecto muy corto, algo flexuoso y sin ramificarse,
terminan en una pequeña arborización que recuerda a una garra y que
fue comparada por Rouget a una placa motora. Estas terminaciones
dendríticas confluyen en los islotes protoplásmicos, donde van a
intervenir en una sinapsis múltiple que ya estudiaremos. Su
superficie es lisa, no presentan contactos sinápticos más que en la
garra y mantienen un diámetro constante en todo su recorrido. Su
ultraestructura presenta neurotúbulos y neurofilamentos. Desde el
punto de vista inmunohistoquímico, los granos son positivos a la
anti-calretinina con cuya técnica, según Yan y Garey (39) aparecen
agrupados en asociaciones de aproximadamente seis unidades. (fig.
10).
El axón nace habitualmente del soma o, en
raras ocasiones de la dendrita más alta, y asciende más o menos
curvadamente hasta la capa molecular. Este trayecto ascendente es al
principio liso, pero luego se torna varicoso (fig.
10). Al llegar a
la capa molecular, el axón se divide en "T", dando dos ramas
contrapuestas que avanzan en sentido longitudinal y que constituyen
las fibras paralelas (F.P.) de Cajal. Estas fibras están
orientadas en el eje longitudinal de la laminilla, de manera que
para verlas en toda su longitud hemos de utilizar cortes también
longitudinales, en los que veremos el perfil del ramaje dendrítico
de las células de Purkinje, imagen que fue comparada por Cajal a un
tendido telefónico. Esto significa que las F.P. atraviesan
perpendicularmente el plano del árbol dendrítico de las células de
Purkinje. Habitualmente existe una relación entre la situación de la
fibra paralela, su grosor y su soma, de suerte que los granos más
profundos son los que poseen el axón más grueso y dan origen a las
F.P. más profundas.
En el curso de su trayecto, las FP hacen
abundantísimas sinapsis "en passant" con todos los tipos celulares
de la capa molecular, pero especialmente con las células de Purkinje
y más concretamente con las espinas dendríticas que son
diferenciaciones específicas para las F.P.. Estas sinapsis contienen
vesículas esféricas y conformación tipo I de Gray, lo que se
corresponde con su carácter activador. Generalmente, el
ensanchamiento de la F.P. envuelve a la cabeza de la espina, pero el
disco sináptico es mucho más pequeño, comprendiendo solamente un
15-20% del total de la superficie. No es infrecuente observar que
sobre una espina se producen sinapsis de dos F.P..
Las fibras paralelas fueron consideradas por
Cajal como el prototipo de las conexiones cruciformes de alto poder
generador pues, según él recorrerían la laminilla de punta a punta,
haciendo sinapsis con todas las células de Purkinje. Sin embargo,
los datos cuantitativos conocidos invalidan la suposición, ya que
una C.P. no posee suficientes espinas para recibir contactos
sinápticos de todas las F.P. que atraviesan su campo dendrítico. El
problema a sido revisado por Fox (8), quien demostró que las F.P. no
recorren la totalidad de la laminilla sino solo una longitud de 2-3
mm. y atraviesan los campos dendríticos de 230-460 C.P., lo que
concuerda con datos electrofisiológicos. A su vez, en este trayecto,
tampoco conectan con todas las CP puesto que en el campo anatómico
de una C.P., que contiene aproximadamente 60.000 espinas, pueden
contarse hasta 200-300.000 FP. Puesto que está claro que las únicas
sinapsis F.P.-C.P. se establecen sobre las espinas, se deduce que
solo unas cuantas fibras contactan con la C.P. En resumen, aunque
las F.P. atraviesan los campos dendríticos de 230-460 C.P., solo
sinaptan con una de cada cinco y cada F.P. contacta con 46-92 C.P.
Esto tiene su importancia para la comprensión de la arquitectura
funcional.
GRANDES CÉLULAS ESTRELLADAS.
Las grandes células estrelladas son un
elemento sobre el que existe una gran confusión terminológica, que
ha sido revisado muy profundamente con M.O. por Cajal (4) y a la luz
de más recientes aportaciones por Mugnaini (1974) (21). En la
siguiente exposición intentaremos aclarar y simplificar conceptos en
la medida de lo posible, atendiendo solamente a aquellos datos con
coherencia morfofuncional.
Para ello, consideraremos, de momento, como
grandes estrelladas a toda neurona distinta a los granos y ubicada
en esta capa. Con este criterio, podemos distinguir de momento las
siguientes variedades:
a) Células de soma grande, dendritas
rectilíneas dirigidas a la molecular y axón corto y muy
ramificado en la capa de los granos. A estas células que tienen
un significado funcional importante las llamaremos células de
Golgi.
b) Elementos idénticos a los anteriores
pero en situación ectópica (capa molecular o sustancia blanca),
denominadas por Cajal células desplazadas. Este tipo es idéntico
al anterior, pero se trata de elementos aberrantes y por tanto
muy infrecuentes y con escasa significación funcional.
c) Células de soma fusiforme grande y
dendritas opositopolares rectilíneas o en abanico, que se
extienden inmediatamente por debajo de los somas de las células
de Purkinje. Este tipo fue denominado por Cajal "grandes
estrelladas fusiformes", ahora conocidas como células de Lugaro
en atención a la descripción minuciosa que este autor hizo de
ellas.
d) Células de soma grande, con dendritas
como las células de Lugaro o de Golgi, pero cuyo axón se dirige
a la sustancia blanca. Se trata también de elementos aberrantes,
bien pertenecientes a los núcleos grises (Cajal) o bien células
de Purkinje aberrantes (Estable, Eccles, Ito, Szentagothai). Son
elementos de escasa significación que no estudiaremos.
De todo lo anterior podemos concluir que
dentro de lo que Cajal designó como grandes estrelladas, hay dos
tipos de valor funcional e histológico definido, las células de
Golgi y las de Lugaro.
CÉLULAS DE GOLGI:
Son elementos que
tienen el soma estrellado y están situados a todas las alturas en la
capa de los granos. Contienen grumos de Nissl y abundantes
neurofibrillas, así como un complejo de Golgi y R.E.L. casi tan
ricos como en las C.P.; sin embargo las cisternas hipolemnales son
bastante menos frecuentes. El núcleo es a veces escotado, de
cromatina laxa y con un nucleolo prominente y que suele ocupar una
posición excéntrica. Sobre la superficie del soma pueden encontrarse
botones sinápticos pequeños, correspondientes a colaterales de
Purkinje y que presentan la misma morfología descrita para la
terminación principal, con vesículas aplanadas que estructuran
sinapsis tipo II de Gray. Sobre el soma se pueden observar otras
sinapsis más grandes que describiremos más adelante y que
corresponden a terminales de fibras extrínsecas al cerebelo. Desde
un punto de vista inmunohistoquímico, según Sahin y Hockfield (34),
estas células son positivas al anticuerpo Rat-303 en el gato,
mientras que Négyessy y cols., 1997 (26), observan su positividad
para el mGluR5.
Las dendritas nacen de 4-5 gruesos troncos
principales y después de un breve trayecto horizontal o descendente
se dicotomizan e incurvan para alcanzar la capa molecular. Son
expansiones casi rectilíneas, poco ramificadas y casi sin
varicosidades, aunque en su trayecto por la capa de los granos,
presentan unas excrecencias groseras que les prestan un aspecto
villoso (21). No se trata de espinas, sino simplemente de
irregularidades del contorno dendrítico, que representan
dispositivos postsinápticos para fibras extrínsecas. En los espacios
que dejan libres estas excrecencias, se encuentran botones
sinápticos pertenecientes a colaterales recurrentes de Purkinje
similares a los que encontrábamos en el soma y sinapsis "en passant"
formadas por el segmento ascendente de los axones de los granos con
contactos tipo I de vesículas redondas en la presinapsis.
La ultraestructura de las dendritas es similar
a la del soma, aunque los organoides van disminuyendo a medida que
nos alejamos y en la capa molecular solo contienen haces de
neurotúbulos y algo de R.E.L.. También a este nivel pueden verse
abundantes contactos sinápticos de las fibras paralelas y de
elementos propios de la capa plexiforme. El campo dendrítico de la
célula de Golgi está dispuesto, a diferencia del de la célula de
Purkinje, en los tres planos espaciales y comprende un amplio
territorio; sus dendritas abarcan un área dentro de la cual se
albergan 16-25 células de Purkinje, sin que exista apenas
superposición de los campos.
El axón de estas células es su elemento más
característico. Nace del soma o de uno de los troncos dendríticos
principales y presenta un segmento inicial muy breve (4-5 µm.). A
partir de este momento, se divide formando un plexo intrincadísimo;
tanto que estas células son el ejemplo típico de neuronas de axón
corto. Las ramificaciones son siempre en ángulo recto y se extienden
por todo el espesor de la capa de los granos subyacente al soma. Las
ramas terminales del axón se dirigen y acaban en los islotes
protoplásmicos, donde en unión de las dendritas de los granos y de
fibras extrínsecas van a modelar una sinapsis compleja.
El patrón de arborización axónico fue muy bien
estudiado por Cajal, quien aisló tres tipos básicos que luego han
resultado tener una correspondencia funcional perfecta. En el primer
tipo, el plexo axónico cubriría un campo mas o menos equivalente al
de las dendritas; en el segundo tipo, el axón se extendería mucho
más pero sin salir de la laminilla, en tanto que en el tercer tipo,
del segmento inicial se originan dos plexos, uno en la propia
laminilla y otro en la vecina.
CÉLULAS DE LUGARO:
Son elementos grandes, cuyo soma suele
encontrarse inmediatamente por debajo de la hilera de somas de
Purkinje, existiendo confusión en cuanto a su naturaleza. En 1894,
Lugaro describió una célula situada por debajo de las C.P. con
dendritas largas y opositopolares y un axón que, tras un corto
trayecto, se resolvería en un plexo inmediatamente por encima de los
somas de las C.P.. Este tipo de arborización axónica no fue
encontrado por Cajal, quien sí pudo identificar, sin embargo, el
soma y dendritas de las células descritas por Lugaro, denominandolas
células fusiformes horizontales. En los trabajos de Cajal, las
células fusiformes horizontales tendrían dos tipos de organización
axónica: en una de ellas el axón ingresaría en la sustancia blanca y
en la otra se resolvería dentro de la capa de los granos en un plexo
idéntico al de las células de Golgi. Los resultados de Cajal fueron
reinterpretados posteriormente, considerándose que las fusiformes
horizontales con axón a la sustancia blanca serían C.P. aberrantes,
en tanto que las de axón corto no serían sino células de Golgi cuyas
dendritas no ascenderían a la capa molecular. Estas consideraciones
fueron generalmente admitidas hasta que en 1959 Fox
(7) resucitó el
problema al encontrar en el mono células de dendritas opositopolares
cuyo axón se ramificaba, exactamente como dijo Lugaro, por encima
del soma de las células de Purkinje. Posteriormente Palay (27)
comprobó los resultados de Fox aunque también ha encontrado células
cuyo axón parece ir a la sustancia blanca. Por lo tanto, en el
momento actual se puede pensar en que quizá haya varios tipos de
estas células grandes de dendritas opositopolares, algunas con axón
corto y otras con axón largo. En la pasada década (1996), Lainé y
Axelrad (15) vuelven al estudio de este tipo neuronal y lo describen
con el árbol dendrítico ya conocido anteriormente, mientras que el
axón se bifurca en un amplio plexo arrosariado que abarca la zona
superior de la capa granulosa y la casi totalidad de la molecular,
dispuestos en el plano sagital. Por otra parte Sahin y Hockfield
(34), ya habían demostrado en 1990 la caracterización de estas
células en el gato al encontrarlas, a diferencia de todas las demás,
positivas a los anticuerpos Cat-301 y Cat-304, mientras que Négyessy
y cols., 1997 (26), observan su positividad para el mGluR5 que
también mostraban las células de Golgi.
El confusionismo se debe a la extraordinaria
dificultad para impregnar el axón, lo que a su vez conlleva que la
sinaptología sea mal conocida, puesto que no es facil identificarlo
ni con el M.E. ni con las modernas técnicas de inmunohistoquimia.
Sin embargo interesa destacar que son elementos de presencia
constante, cuyas dendritas se extienden en el plano transversal,
cubriendo un campo que alberga 1-2 hileras completas de células de
Purkinje. Esta disposición es idéntica a la de los grandes elementos
de asociación de la VII capa de la retina. Aunque aún no se puede
asegurar, es posible que juegen un papel modulador y que la
dificultad de impregnar su axón pueda ser debida a que se trate de
células de tipo amacrino, de transmisión bidireccional, que actúen
directamente sobre los axones de las C.P. que salen a este nivel. En
suma, son células de momento mal conocidas pero que posiblemente
jueguen un papel importante en la elaboración de la respuesta
cerebelosa.
CÉLULAS MONODENDRÍTICAS EN PENACHO:
Recientemente se ha descrito un nuevo tipo
celular, cuyo significado funcional no está claro, que se sitúa en
la capa de los granos de la corteza cerebelosa de los mamíferos. Se
trata de la célula monodendrítica o monopolar en penacho,
descubierta por Muñóz en 1990 (25) y descrita posteriormente por Braak y Braak
(3), Mugnaini y Floris
(23) y Yan y Garey (39).
Presenta un soma esférico y un solo tronco dendrítico que termina en
una corta arborización en penacho y parece tratarse de una célula de
asociación.
Arriba
CAPA MOLECULAR
Como ya hemos apuntado, esta capa es una banda
parvicelular de aproximadamente 300-400 µm. de espesor que se
extiende entre la hilera de los somas de la células de Purkinje y la
píamadre. Fundamentalmente está constituida por un plexo muy tupido
de axones y dendritas, entre el que se distribuyen como salpicados
los somas de los únicos elementos neuronales que residen en ella y
que son las células estrelladas pequeñas, que pueden ser
superficiales (células estrelladas de la terminología actual) y
profundas (células en cesta), situadas estas inmediatamente por
encima de los somas de las C.P..
CÉLULAS EN CESTA:
Fueron entrevistas por Golgi y descritas por
Cajal (4), quien las denominó "pequeñas estrelladas profundas",
aunque se instauró, por parecer más oportuno, el nombre de células
en cesta (Korbzellen) propuesto por Kölliker en atención al modo de
terminación de sus axones.
Son elementos cuyo soma está justo por encima
de la hilera de somas de Purkinje, presenta morfología triangular o
estrellada y un diámetro promedio de 10-20 µm. Tienen un núcleo
lobulado y excéntrico y su citoplasma posee escasos organoides
concentrados en el polo que deja libre el núcleo; los grumos de
Nissl son pequeños y escasos, el complejo de Golgi y el R.E.L. son
poco prominentes y puede presentar alguna cisterna hipolemnal
(fig.
13).
Las dendritas, que se ramifican en el plano
transversal, pueden ser cortas y descendentes, pero lo habitual es
que asciendan hasta el tercio superior de la capa plexiforme. Son
relativamente rectilíneas y poco dicotomizadas, presentando espinas
aunque más escasas y groseras que las de las C.P.. Contienen
abundantes neurotúbulos y neurofilamentos, R.E.L. incluso en sus
porciones distales, mitocondrias alargadas y algo de R.E.R. y
complejo de Golgi en los troncos principales.
Sobre el soma y las dendritas de las células
en cesta terminan importantes aferencias. Así son abundantes las
sinapsis "en passant" de las fibras paralelas, las cuales asientan
sobre las espinas y son tipo I de Gray con discos sinápticos
extensos. Tanto sobre el soma como sobre las dendritas pueden
identificarse otros botones sinápticos, tipo II de Gray que
corresponden a axones de otras células en cesta y de estrelladas
superficiales, así como colaterales del plexo supragangliónico de
las C.P.
El axón es amielínico, nace del soma como una
expansión muy fina que discurre para sagitalmente sobre el campo
dendrítico de 1-2 células de Purkinje. Esta primera porción, que
presenta características de segmento inicial, termina bruscamente
con un aumento de calibre que ya no variará y a partir del cual
comienza a dar colaterales. Estas colaterales, ricas en
neurofibrillas y de baja densidad electrónica, siguen un curso en el
plano longitudinal, dando ramas ascendentes y descendentes. Las
ascendentes, que son generalmente más cortas, llegan hasta la zona
media de la capa molecular y en este camino sinaptan, en estructuras
tipo II de gray, con dendritas lisas de las C.P., dendritas de otras
células en cesta y posiblemente con dendritas de las células de
Golgi.
Las colaterales descendentes, forman un
dispositivo en el que hizo Cajal la observación inicial que le
permitió enunciar la teoría neuronal. Se trata de una sinapsis
compleja en la que las colaterales descendentes abrazan por completo
el soma de la célula de Purkinje (fig. 3), confluyendo sus extremos
en la base del soma donde forman un pincel que rodea al axón de la
célula de Purkinje. El análisis E.M. ha venido a añadir nuevas
peculiaridades de esta singular sinapsis. Los principales estudios
fueron efectuados inicialmente por Hamori y Szentopothai
(9), siendo
posteriormente corregidos por Sotelo (38), Llinas
(19), Palay (27), Mugnaini
(20), etc. quienes describen el soma completamente rodeado
por células gliales, por fuera de las cuales se deslizan las ramas
descendentes de la cesta, las cuales taladran la envoltura glial y
sinaptan con el soma, dirigiéndose luego al segmento inicial axónico,
donde conforman el pinceaux. En este lugar existe una tremenda
convergencia, observándose multitud de axones preterminales
dispuestos de forma concéntrica alrededor del delgado segmento
inicial axónico de la C.P.. Entre estos axones se forman uniones
comunicantes, pero curiosamente, las sinapsis químicas sobre el
propio segmento inicial son muy escasas y según opinión de Mugnaini,
en todo el pinceaux solo se forman 1-2 sinapsis sobre el segmento
inicial. En el resto de su trayecto, este aparece en aposición con
los axones del pinceaux, existiendo entre ambas estructuras un
espacio intercelular bien definido, no identificándose ningún tipo
de diferenciaciones de membrana.
En cuanto a la disposición geométrica del
campo axónico de las células en cesta hay que apuntar que, al
contrario que en otras células del cerebelo, existe una notable
superposición en los campos axónicos de las cestas. Según distintos
cálculos cada célula en cesta actúa a través de sus colaterales
axónicas descendentes sobre un número determinado de C.P. (216 en el
mono, Fox (8), 60 en el gato, Palkovits
(30)), que además no están
repartidas de manera anárquica sino siguiendo un patrón más o menos
rectangular (12x20 en el mono, 6x10 en el gato, 3x3 en la rata).
Estos campos rectangulares están muy superpuestos, de forma que en
una célula de Purkinje intervienen colaterales de distintas células
en cesta.
CÉLULAS ESTRELLADAS:
Se trata de células entrevistas por Fusari y
luego estudiadas a fondo por Cajal, que las clasifica en dos grandes
clases, de axón corto y de axón largo, aunque esta clasificación fue
posteriormente criticada, más utilizando criterios histofuncionales
que morfológicos.
Son neuronas de soma pequeño (5-8 µm.),
poligonal o estrellado, que alberga un núcleo de cromatina laxa y un
citoplasma con pocos organoides. Las dendritas, que se ramifican en
el plano transversal, parten de cinco o seis troncos dendríticos
principales y generan un plexo circunscrito de ramas varicosas,
provistas de espinas. Este plexo dendrítico, habitualmente comprende
solo una parte del campo de 1-2 C.P.. Sobre soma y dendritas se
producen sinapsis "en passant" tipo I formadas por las fibras
paralelas y sinapsis tipo II correspondientes a axones de otras
estrelladas y colaterales ascendentes de los axones de células en
cesta. El axón surge de un cono axónico bien definido y después de
un segmento inicial de 5-6 µm. de longitud, se resuelve en un plexo
circunscrito, aunque desplazado del soma y orientado verticalmente,
extendiéndose por los campos dendríticos de las 4-8 células de
Purkinje circundantes. Las colaterales axónicas acaban formando
contactos tipo II con las dendritas y somas de las C.P., existiendo
también contactos del mismo tipo con las dendritas de otras
estrelladas, células en cesta y células de golgi.
Además existen otras, más grandes que las
anteriores, presentando su soma una estructura idéntica al de las
células en cesta, del que parten 5-6 expansiones que se dirigen de
forma rectilínea hacia la zona más superficial de la capa plexiforme,
presentando algunas bifurcaciones y espinas groseras como las de las
células en cesta. El axón, después de un segmento inicial de 9-10 µm.
de longitud, se engruesa y corre en el plano parasagital, dando
colaterales transversales de las que a su vez, emergen ramas
ascendentes y descendentes. Las ascendentes hacen sinapsis tipo II
con dendritas de las cestas, estrelladas y Golgi y más
frecuentemente con dendritas de las células de Purkinje. Las
colaterales descendentes de las más profundas pueden participar en
la formación de la cesta, aunque no está claro si llegan a formar
parte del pinceaux. Salvo este detalle, estas células son idénticas
a células en cesta desplazadas y así las consideró Cajal (4) y Palkovits (29), aunque más recientemente Palay (27) criticó esta
consideración. Posiblemente sean células diferentes a cestas
desplazadas, pero esta diferencia es en base a su posible
significación fisiológica más que a su morfología, pues como el
mismo Palay confiesa, solo en algunos casos son diferentes de las
células en cesta.
Arriba
FIBRAS EXTRÍNSECAS.
Sabemos que las eferencias la corteza son los
axones de las células de Purkinje, pero nos resta conocer de donde
se nutre toda la circuitería de la corteza, es decir las aferencias.
Existen dos tipos de fibras aferentes que llegan a la corteza
cerebelosa: Las fibras musgosas y las fibras trepadoras.
Ambas ingresan en la corteza desde la sustancia blanca y van a
conectar con elementos neuronales de la misma.
FIBRAS MUSGOSAS:
Son gruesas fibras mielínicas que penetran en
el cerebelo por cualquiera de los pedúnculos y que después de
dividirse profusamente en la sustancia blanca, dan ramas que
terminan en los núcleos grises, formando con las neuronas del núcleo
sinapsis que se producen en la terminación de la fibra, que se
indenta en el citoplasma neuronal formando una estructura redondeada
parecida a una castaña (de ahí la denominación de sinapsis "en
marron" de los franceses) donde se sitúa el aparato presináptico de
un contacto sináptico tipo I de Gray. Otras ramas abordan la corteza
penetrando en la capa de los granos, donde siguen un curso tortuoso,
dando multitud de colaterales. Durante su trayecto por la granulosa,
las fibras musgosas presentan "unos abultamientos nudosos que se
diría están constiutidos por un acúmulo irregular de plata
precipitada. Estas varicosidades son verdaderas arborizaciones
cortas y varicosas que aparecen en ciertos parajes de la fibra a la
manera de un musgo o maleza de revestimiento. En numerosas
ocasiones, esta arborización está sostenida por un tallo corto y
delgado que le presta el aspecto de una flor" (Cajal, 1888). Estas
eflorescencias, que se conocen como rosaceas o rosetas, pueden estar
localizadas no solo en el curso de la fibra, sino también en su
terminación y en las bifurcaciones y como ya postuló Cajal,
representan dispositivos de articulación sináptica que se disponen
en aquellas estructuras acidófilas, denominadas islotes
protoplasmicos, donde también confluían las dendritas de los granos
y las terminaciones axónicas de las células de Golgi. Estos tres
elementos van a constituir una estructura sináptica compleja que se
conoce desde Held y fue estudiada ultraestructuralmente por primera
vez por Palkovits (29,31), quien confirmó las ideas de Cajal,
denominada: glomérulo cerebeloso (fig. 14).
Las fibras musgosas son gruesas, con
abundantes neurotúbulos, neurofilamentos y mitocondrias. Presentan
una gruesa vaina de mielina, en cuyos nodos de Ranvier residen las
eflorescencias descritas por Cajal; en el contorno de estos
ensanchamientos irregulares de la fibra se indentan las
terminaciones dendríticas de los granos, mientras que el centro de
la rosacea suele estar ocupado por neurofilamentos y mitocondrias,
bordeando a las cuales aparecen abundantísimas vesículas esféricas
configurando la presinapsis que forma multiutd de contactos tipo I
con las dendritas de los granos. El tercer elemento del glomérulo
son las terminaciones del plexo axónico de las células de Golgi. Se
trata de expansiones electroclaras que forman sinapsis tipo II con
las dendritas de los granos, en situación contrapuesta al disco
sináptico que forman la fibra musgosa y la dendrita. En el glomérulo
no existen contactos sinápticos entre la rosacea y los axones de la
célula de Golgi, pese a que en algunos lugares puedan estar
anatómicamente próximos.
Con M.E. se ha comprobado (31) la observación
previa de Cajal de que existen sinapsis entre las rosaceas y el soma
y/o dendritas de las células de Golgi. Estas sinapsis con las
dendritas poseen numerosos discos sinápticos de gran extensión, no
forman glomérulo y la especialización sináptica se sitúa solo en la
cara que mira a la dendrita. En el caso de los contactos con el soma
neuronal, presentan una disposición "en marron" con la estructura de
la rosacea incluida en una indentación del soma, lo que les presta
una morfología redondeada. Estas sinapsis son de tipo I y su
importancia reside en que al contener multitud de discos sinápticos,
su poder activador es muy elevado.
FIBRAS TREPADORAS:
Son fibras de menor diámetro que las musgosas,
descritas por Cajal en 1888, que penetran a través de los pedúnculos
cerebelosos y tras dar colaterales para todos los núcleos grises,
donde contactan con la neuronas del núcleo efectuando sinapsis "en
marron" de tipo I, alcanzan la corteza y se ramifican sobre todo en
la capa molecular. Atraviesan la capa granulosa de forma casi
rectilínea, con escasas varicosidades y dando únicamente 1-2
colaterales. Al alcanzar el nivel del soma de las células de
Purkinje, la fibra pierde la mielina y da colaterales ascendentes
que trepan, a modo de lianas, por los troncos dendríticos
principales de las C.P.. En el curso de este ascenso, dan lugar a
abundantes ramas delgadas que, tras un breve y flexuoso trayecto,
terminan en un botón. Tanto estos botones como las varicosidades del
plexo principal, corresponden a dispositivos sinápticos sobre el
árbol dendrítico de las C.P. (fig. 15).
Observadas mediante M.E., las fibras
trepadoras en su último trayecto son finas y amielínicas, con
algunos neurofilamentos, pocas mitocondrias y abundantes sinapsis
"en passant" tipo I con las dendritas de las C.P.. La identificación
elecromicroscópica de los terminales de estas fibras fue obra del
español Larramendi (16), quien demostró que sinaptan sobre ciertas
excrecencias de los troncos dendríticos principales que ya
describimos anteriormente. Con este hallazgo se vino a completar la
arquitectura sinaptológica de las células de Purkinje.
Por otro lado, la identificación E.M. de las
trepadoras con sus botones muy densos y atiborrados de vesículas
redondas, ha permitido comprobar, como ya dijera Cajal, la
existencia de sinapsis entre estas fibras y las dendritas de las
células estrelladas y las células en cesta. El destino de las
colaterales que permanecían en la capa de los granos son las
dendritas de las células de Golgi y el soma de las mismas, con el
que estructuran un aparato sináptico similar al que provocaban las
fibras musgosas (27), esto es, una estructura redondeada incluida en
la indentación del soma con múltiples discos sinápticos tipo I.
Arriba
NEUROGLIA DEL CEREBELO
En la sustancia blanca existen astrocitos
fibrosos y oligodendrocitos sin características especiales. En la
capa de los granos se pueden observar astrocitos protoplásmicos que
no aíslan todas las neuronas y que parecen formar circulos alrededor
de los glomérulos. En la capa molecular se sitúan unas células
gliales denominadas de Bergmann que presentan un cuerpo
irregular localizado al lado de los somas de las C.P. y que emiten
de una a tres expansiones muy varicosas que atraviesan la capa
molecular y se adosan, mediante ensanchamientos que forman la
limitante de Cajal, en la piamadre. Existen otras células similares
a las anteriores cuyos cuerpos celulares se sitúan en posiciones más
altas en la capa molecular y cuyas expansiones no alcanzan la
piamadre que se denominan células de Fañanás. En realidad,
tanto las células de Bergmann como las de Fañanás son variedades de
astrocitos protoplásmicos, adaptados a la morfología en candelabro
de la C.P. y que no presentan ninguna peculiaridad ultraestructural
(fig. 16), expresando ambas positividad para el anticuerpo de la
proteína ácida gliofibrilar. Se ha descrito también la presencia de
oligodendrocitos en la capa molecular pero no en la de los granos.
HISTOFISIOLOGÍA
Como hemos visto hasta ahora, el cerebelo es
quizá el territorio del S.N.C. que presenta la estructura más
geométrica y mejor conocida, siendo posible establecer, incluso
desde bases puramente morfológicas, una serie de correlaciones
funcionales. Está bien comprobada la existencia de sinapsis
musgosas-granos, fibras paralelas-Células de Purkinje, fibras
paralelas-células en cesta, células en cesta-células de Purkinje,
etc., pero, en contra de lo que pensaba Cajal, no todas las sinapsis
son activadoras. Debido a que las neuronas de asociación tienen
actividad inhibidora no parece sostenerse la teoría de que la misión
de estas es la de repartir y ampliar el estímulo a campos más o
menos alejados. Actualmente se sostiene que la misión de las
neuronas de asociación o intercalares, más que repartir el estímulo
por las áreas cercanas es precisamente la contraria, impedir la
difusión incontrolada de los impulsos
En general, se puede describir un circuito
principal que penetraría en la corteza mediante las fibras musgosas
y que a través de los contactos con los granos en el glomérulo,
modulados o inhibidos por la célula de Golgi, y los de las fibras
paralelas con la C.P. conseguiría salir de la corteza mediante los
axones de las C.P.. A su vez otro circuito que amplificaría el
anterior gracias a su contacto con las C.P., sería el que penetra en
la corteza por las fibras trepadoras.
Estos circuitos principales, se ven modulados
por las neuronas de asociación que producen contactos inhibidores,
tanto en el glomérulo (células de Golgi) como en las dendritas de
las C.P. (Células estrelladas), en el segmento inicial del axón de
la C.P. (Células en cesta) o la función, de momento más oscura que
puedan tener las células de Lugaro o las monodendríticas en
ramillete.
A su vez, la única vía eferente de la corteza
está representada por los axones de las células de Purkinje que
formarán sinapsis inhibidoras que modulan el contacto activador que
se produce entre las fibras musgosas y trepadoras y las neuronas de
los núcleos grises del cerebelo (fig. 17).
Arriba
BIBLIOGRAFÍA
1.- Angaut, P. Y Sotelo, C. (1975): Diversity
of mossy fibres in the cerebellar cortex in relation to different
afferent systems: an experimental electrón microscopic study in the
cat.
2.- Bloedel, J. R. y Courville, J.
(1981): Cerebellar afferent systems. En: J.M. Brookhart, V.B.
Mountcastle and V.B. Brooks (Eds.), Handbook of Fhysiology, Vol. 2,
Am. Physiol. Soc., Bethesda. Pp 735-829.
3.- Braak, E. y Braak, H.
(1993): The new
monodendritic neuronal type within the adult human cerebellar
granule cell layer shows calretinin-immunoreactivity. Neurosci. Lett.
154(1-2): 199-202.
[volver al texto]
4.- Cajal, S.R.
(1904): Textura del sistema nervioso del hombre y los vertebrados.
Ed. Nicolás Moya.
Madrid.
[volver al texto]
5.- Díaz Flores, L.; Gayoso, M.J.; Sanchez, G;
Aneiros, J.; Aguilar, D.; Ortíz, G.; Linares, J.; Caballero, T.;
Spreáfico, J.M.; Garrido, M.; Alonso, J.; del Moral, R.G.; Martos,
S. y Lucas, A.R. (1977): Neurohistología. Granada.
6.- Eccles, J.C.; Ito, M. y Szentagothai, J.
(1967): The cerebellum as a neuronal machine. Ed. Springer-Verlag.
Berlin.
[volver al texto]
7.- Fox, C.A. (1959): The intermediate cells
of Lugaro in the cerebellar cortex of the monkey. J. Comp. Neurol..
112, 39-53.
[volver al texto]
8.- Fox, C.A., Hillman, D.E., Siegesmund, K.A. y
Dutta, C.R. (1967): The primate cerebellar cortex: a Golgi and
electrón microscopic study. Prog. Brain Res. 25: 174-225.
[volver al texto]
9.- Hamori, J. y Szentagothai, J. (1966):
Identification under the electrón microscope of climbing fibers and
their synaptic contacts. Exp. Brain. Res. 1(1): 65-81.
[volver al texto]
10.- Hawkes, R y Leclerc, N.
(1989): Purjinje cell axon collateral distributions reflect de
chemical compartmentation of the rat cerellar cortex. Brain Res.
476(2): 279-90.
[volver al texto]
11.- Ito, M. (1968): The neuronal mechanism
of the cerebellar efferent system. Proc. Aust. Assoc. Neurol. 5(1):
13-8.
12.- Ito, M.
(1969): Neurons of cerebellar nuclei. UCLA Forum Med. Sci. 11:
309-27.
13.- Ito, M. Sato, N. Simpson, J.I. y Udo, M.
(1971): Contribution of mossy fiber-granule cell pathway to the
cerebellar-induced delayed inhibition in Deiters neurones. Exp.
Brain Res. 12(3): 223-37.
14.- Ito, M. y Simpson, J.I.
(1971): Discharges in
Purkinje cell axons during climbing fiver activation. Brain Res.
31(1): 215-9.
15.- Laine, J. y Axelrad, H.
(1994): The candelabrum
cell: a new interneuron in the cerebelar cortex. J. Comp. Neurol.
339, 159-173.
[volver al texto]
16.- Larramendi, L.M.H. y Lemkey-Johnston, N.
(1967): Synapses on the Purkinje cell spines in the mouse. An
electromicroscopic study.Brain Res.5(1): 15-30.
[volver al texto]
17.- Larsell, O. y Jansen, J.
(1972): The comparative anatomy and histology of the cerebellum. The
human cerebellum, cerebellar connections, and cerebellar cortex. Ed.
University of Minnesota Press, Minneapolis.
[volver al texto]
18.- Leclerc, N.; Dore,, L.; Parent, A. y
Hawkes, R.
(1990): The compartmentalization of the monkey and the rat
cerebellar cortex: Zebrin I and cytochrome oxidase. Brain Res. 506,
70-78.
19.- Llinás, R. (1982): General discussion:
radical connectivity in the cerebellar cortex; a novel view
regarding the functional organization of the molecular layer. En:
The cerebellum. New vistas. Palay, S.L. y Chan-Palay V. (eds.).
Springer-Verlag. Berlin, Heidelberg, New York. Pp 189-194.
[volver al texto]
20.- Mugnaini E.
(1971): Developmental
aspects of synaptology with special emphasis upon cerebellar cortex.
UCLA Forum. Med. Sci. 14: 141-65.
[volver al texto]
21.- Mugnaini E., Atluri, R.L. y Houk, J.C.
(1974):
Fine structure of granular layer in turtle cerebellum with emphasis
on large glomeruli. J. Neurophysiol. 37(1): 1-29.
[volver al texto]
22.- Mugnaini E., Dahl, A.L.
(1975): Mode of distribution of aminergic fibers in the cerebellar
cortex of the chicken. J. comp.. Neurol. 162(4): 417-32.
23.- Mugnaini E. y Floris, A. (1994): The
unipolar brush cell: a neglected neuron of the mamalian cerebelar
cortex. J. Comp. Neurol. 339, 174-180.
[volver al texto]
24.- Mugnaini E. y Forstronen, P.F.
(1967): Ultrastructural studies on the cerebellar histogenesis. I.
Differentiation of granule cells and development of glomeruli in the
chick embryo. Z. Zellforsch. Mikrosk. Anat. 77(1): 115-43.
25.- Muñóz, D.G.
(1990): Monodendritic neurons: a cell type in the human cerebellar
cortex identified by chromogranin A-like immunoreactivity. Brain
Res. 528, 335-338.
[volver al texto]
26.- Negyessy, L. Vidnyansszky, Z. Jun, R.
Knopfel, T. Gorcs, T.J. y Hamori, J. (1997): Light and electrón
microscopic demonstration of mGluR5 metabotropic glutamate receptor
immunoreactive neuronal elements in the rat cerebellar cortex. J.
comp.. Neurol. 385(4): 641-50.
[volver al texto]
27.- Palay, S.L. y Chan-Palay, V.
(1974): Cerebellar cortex: Citology and organization. Ed. Springer-Verlag.
New York.
[volver al texto]
28.- Palkovits, M. Magyar, P. Szentagothai, J.
(1971): Quantitative Histological análisis of the
cerebellarcortex in the cat. I. Number and arrangement in space of
the Purkinje cells. Brain Res. 32(1): 1-13.
[volver al texto]
29.- Palkovits, M. Magyar, P. Szentagothai, J.
(1971):
Quantitative Histological análisis of the cerebellarcortex in the
cat. II. Cell numbers and densities in the granular layer. Brain
Res. 32(1): 15-30.
[volver al texto]
30.- Palkovits, M. Magyar, P. Szentagothai, J.
(1971):
Quantitative Histological análisis of the cerebellarcortex in the
cat. III. Structural organization of the molacular layer. Brain Res.
34(1): 1-18.
[volver al texto]
31.- Palkovits, M. Magyar, P. Szentagothai, J.
(1972):
Quantitative Histological análisis of the cerebellarcortex in the
cat. IV. Mossy fiber-Purkinje cell numerical transfer. Brain Res.
45(1): 15-29.
[volver al texto]
32.- Pellionisz, A. Szentagothai, J.
(1973): Dynamic single unit simulation of a realistic cerebellar
network model. Brain Res. 49(1): 83-99.
33.- Pellionisz, A. Szentagothai, J.
(1974): Dynamic single unit simulation of a realistic cerebellar
network model. II. Purkinje cell activity within the basic circuit
and modified by inhibitory systems. Brain Res. 68(1): 19-40.
34.- Sahin, M. y Hockfield, S.
(1990): Molecular identification of the Lugaro cell in the cat
cerebellar cortex. J. Comp. Neurol. 301, 575-584.
[volver al texto]
35.- Sotelo, C. (1975): Dendritic
abnormalities of Purkinje cells in the cerebellum of neurologic
mutant mice (weaver and staggerer). Adv. Neurol. 12: 335-51.
36.- Sotelo, C. y Changeux, J.P.
(1974): Transsynaptic degeneration "en cascade" in the cerebellar
cortex of staggerer mutant mice. Brain Res. 67(3): 519-26.
37.- Sotelo, C. y Changeux, J.P.
(1974): Bergmann fibers and granular cell migration in the
cerebellum of homozygous weaver mutant mouse. Brain Res. 77(3):
484-91.
38.- Sotelo, C. yLlinas, R. (1972):
Specialized membrane junctions between neurons in the vertébrate
cerebellar cortex. J. Cell. Biol.. 53(2): 271-89.
[volver al texto]
39.- Yan, X.X. y Garey, L.J.
(1996): Calretinin immunoreactiviti in the monkey and cat cerebellum:
cellular localization and modular distribution. J. Hirnforsch.
37(3): 409-19.
[volver al texto]
Arriba
FIGURAS
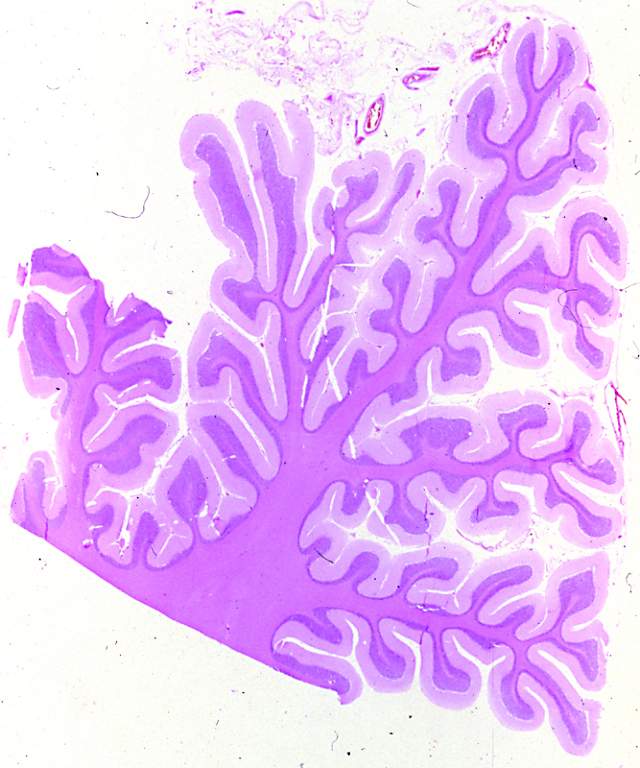
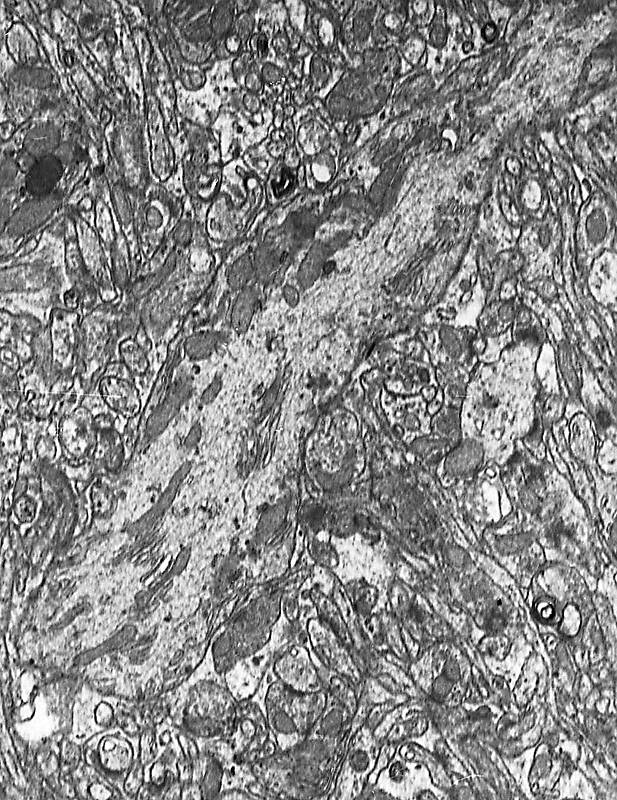
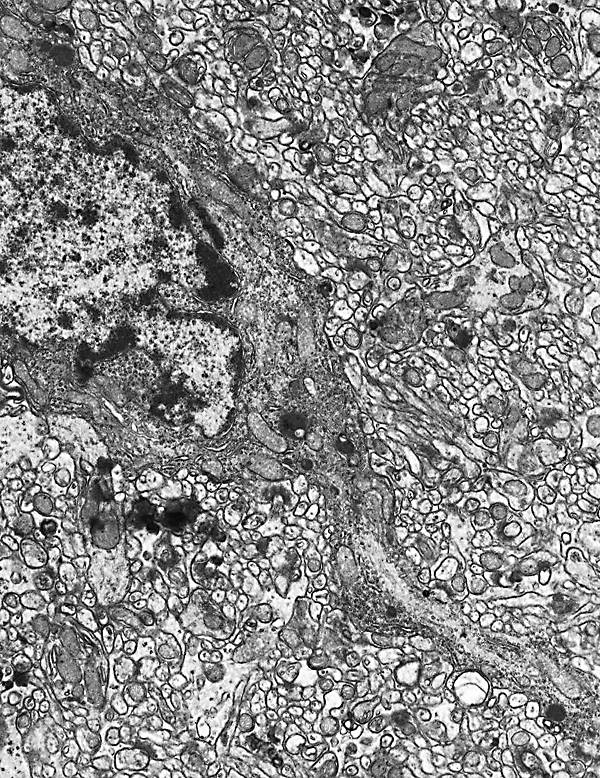
Figura 1
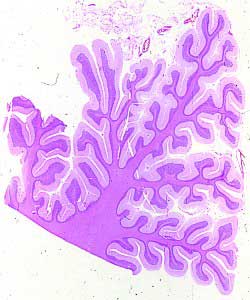
Figura 1.- Microfotografía
panorámica de un corte sagital del cerebelo a pequeño aumento.
Arriba
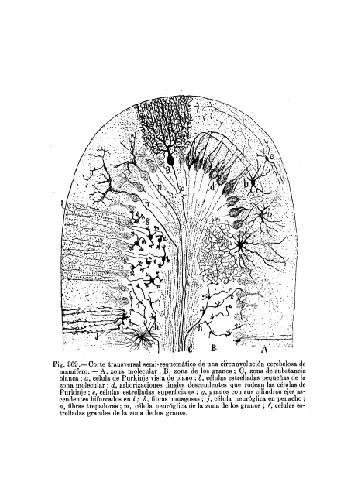
Figura 2
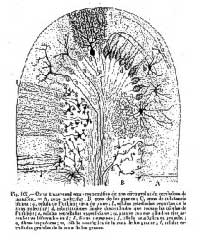
Figura 2.- Esquema de una laminilla
cerebelosa según Cajal.
Arriba
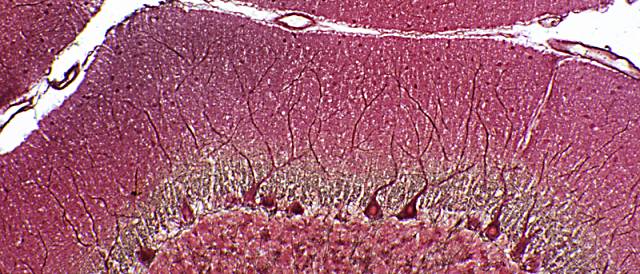
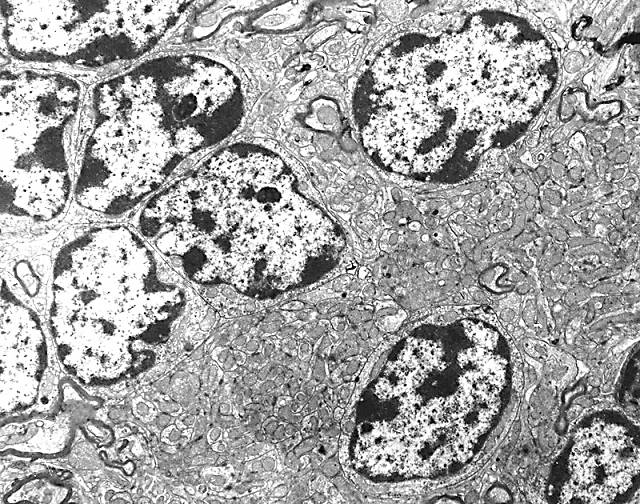
Figura 3
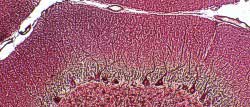
Figura 3.- Microfotografía de la
corteza cerebelosa en la que se aprecian sus dos capas y los somas
de las células de Purkinje en el límite entre las capas, con su
árbol dendrítico. Asimismo se observan, alrededor de los somas de
Purkinje, terminaciones de las células en cesta. (plata reducida de
Cajal).
Arriba
Figura 4
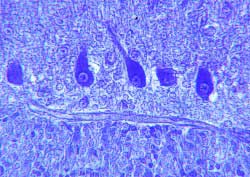
Figura 4.- Somas de las células de
Purkinje mostrando sus núcleos de cromatina laxa y nucleolos
patentes y las granulaciones citoplásmicas. (violeta de cresilo).
Arriba
Figura 5

Figura 5.- Célula
de Purkinje en la que se observa su intrincado árbol dendrítico
(Golgi).
Arriba
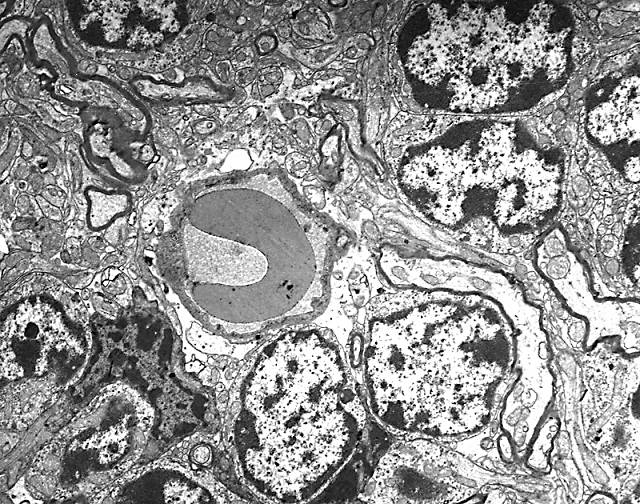
Figura 6
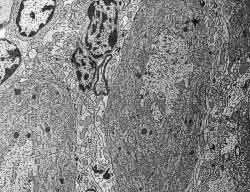
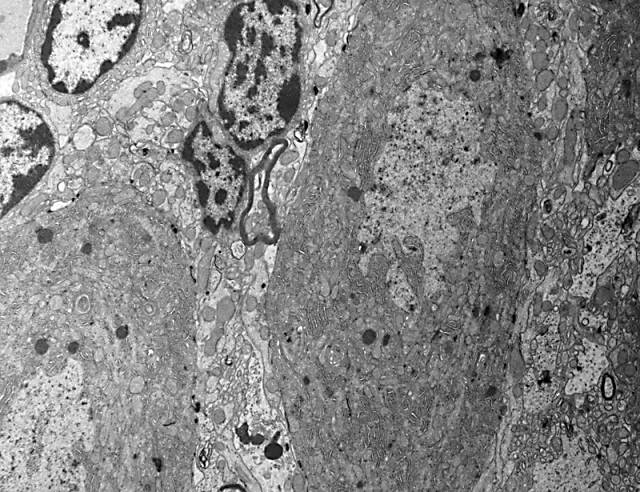
Figura 6.-
Micrografía electrónica en la que se observan los somas de dos
células de Purkinje, células de glia de Bergmann entre ellas y
granos en la esquina superior izquierda.
Arriba
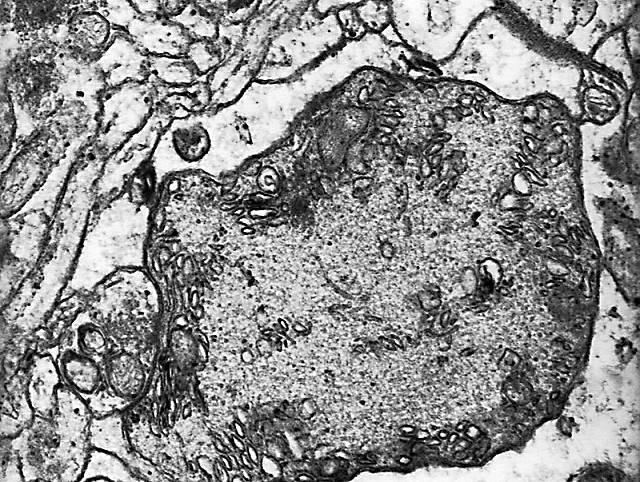
Figura 7
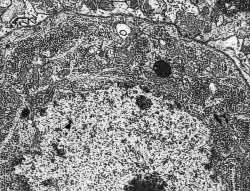
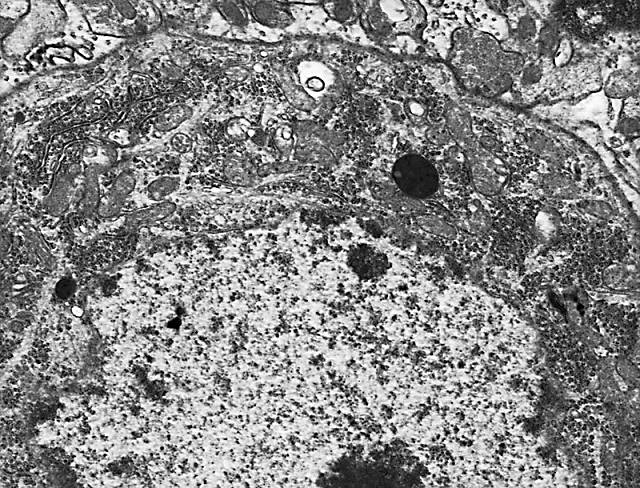
Figura 7.-
Micrografía electrónica del soma de una célula de Purkinge en la que
se muestran algunas características de su núcleo y citoplasma.
Arriba
Figura 8
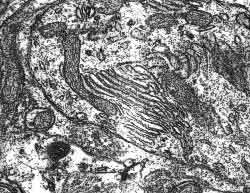
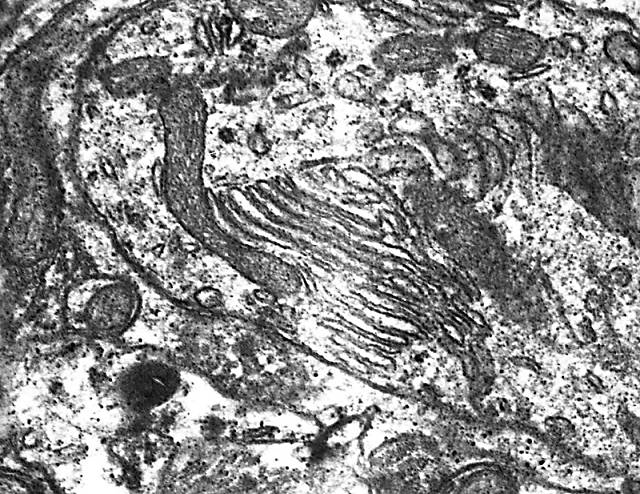
Figura 8.-
Micrografía electrónica de la zona externa del citoplasma de una
célula de Purkinje en la que se observan cisternas hipolemnales
asociadas a una mitocondria.
Arriba
Figura 9
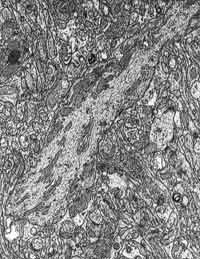
Figura 9.-
Micrografía electrónica de la capa molecular, en la que se observa
una porción de un grueso tronco dendrítico de Purkinje, rodeado de
un intrincadísimo neuropilo.
Arriba
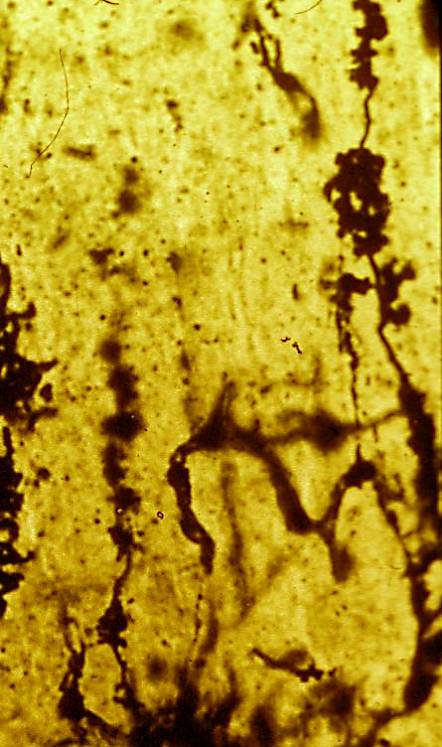
Figura 10
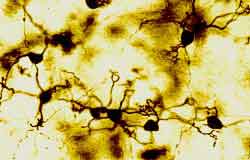
Figura 10.-
Micrografía de la granulosa en la que se muestra el soma de los
granos con sus prolongaciones dendríticas terminadas "en garra" y el
comienzo de sus axones (Golgi).
Arriba
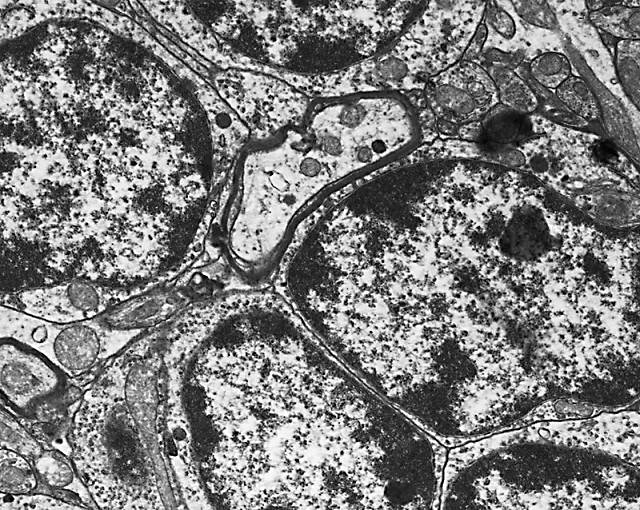
Figura 11
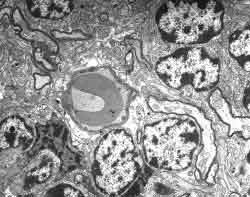
Figura 11.-
Micrografía electrónica de la granulosa en la que se observan los
somas de los granos (mitad derecha), un núcleo de célula de glia y
fibras extrínsecas (cuadrante inferior derecho).
Arriba
Figura 12
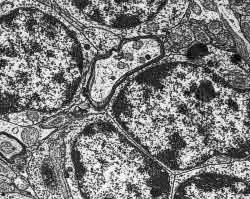
Figura 12.-
Micrografía electrónica que muestra el soma de los granos.
Arriba
Figura 13
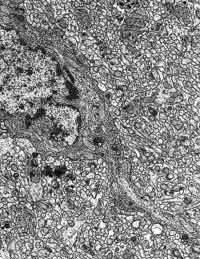
Figura 13.-
Micrografía electrónica de la molecular en la que se observa el soma
y una proyección dendrítica de una neurona estrellada, rodeado de
neuropilo.
Arriba
Figura 14
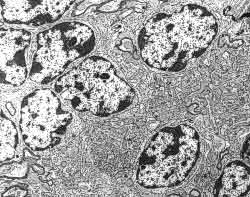
Figura 14.-
Micrografía electrónica de la granular en la que, entre los somas de
los granos se observa un "islote protoplásmico" en el que se
articula el glomérulo cerebeloso.
Arriba
Figura 15
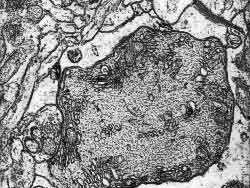
Figura 15.-
Micrografía electrónica de un corte transversal de un tronco
dendrítico de Purkinje, asociado a una fibra trepadora que articula
una sinapsis (cuadrante inferior izquierdo).
Arriba
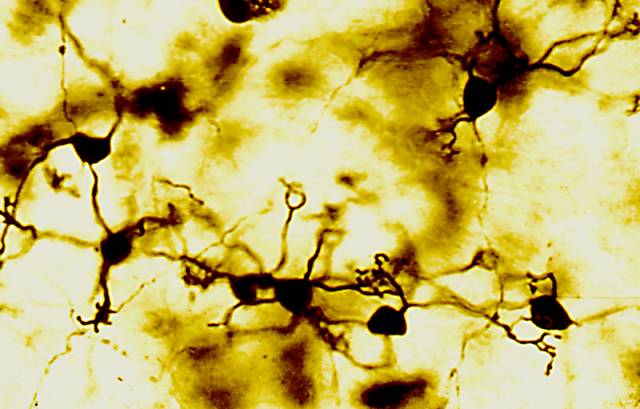
Figura 16
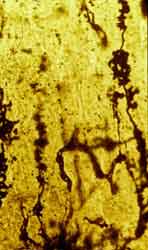
Figura 16.-
Micrografía de la molecular, en la que se muestran representantes de
la glia de Bergmann y Fañanás (Golgi).
Arriba
Figura 17
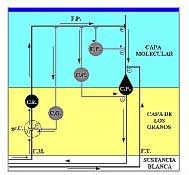
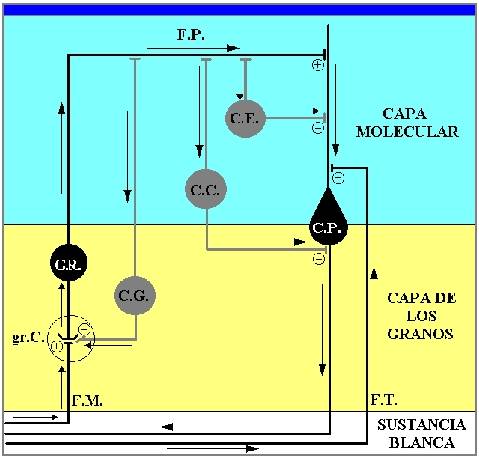
Figura 17.-
Esquema simplificado de los circuitos principales en la corteza
cerebelosa: En negro el circuito excitador. En gris los
circuitos moduladores. C.P.: célula de Purkinje. G.R.:
grano. C.G.: célula de Golgi. C.C.: célula en cesta.
C.E.: célula estrellada. gr. C: glomérulo cerebeloso.
F.M.: fibra musgosa. F.T.: fibra trepadora. F.P.:
fibra paralela. Flechas: sentido del impulso. +:
sinapsis excitadoras. -: sinapsis inhibidoras.
Arriba
--------------------------------------------------------------------------------
|
|
|