1 Database Institute for Chemical Research, Kyoto University. OMIM 104221 Contributors: Victor A. McKusick. Paul J. Converse. Copyright © 1966-2004 Johns Hopkins University.
2 GeneCard for gene ADRA1A - Weizmann Institute of Science
GC08M026627 3 EMBL Bioinformatic Harvester
Q96RE8 - Adrenergic receptor alpha-1a 4 Mouse Genome Informatics - The Jackson Laboratory
MGI:104773 5 Rat Genome Database - Medical College of Wisconsin.
RGD ID: 2055 6 CancerGene - Centre National de la Recherche Scientifique, Institute Gustave-Roussy. Laboratoire de Génétique Oncologique, Groupe de Bioinformatique
CG_ID 2283 7 Hirasawa, A.; Horie, K.; Tanaka, T.; Takagaki, K.; Murai, M.; Yano, J.; Tsujimoto, G. Cloning, functional expression and tissue distribution of human cDNA for the alpha-1C-adrenergic receptor. Biochem. Biophys. Res. Commun. 195: 902-909, 1993.
PubMed ID : 8396931 8 Hirasawa, A.; Shibata, K.; Horie, K.; Takei, Y.; Obika, K.; Tanaka, T.; Muramoto, N.; Takagaki, K.; Yano, J.; Tsujimoto, G. Cloning, functional expression and tissue distribution of human alpha-1C-adrenoceptor splice variants. FEBS Lett. 363: 256-260, 1995.
PubMed ID : 7737411 9 Weinberg, D. H.; Trivedi, P.; Tan, C. P.; Mitra, S.; Perkins-Barrow, A.; Borkowski, D.; Strader, C. D.; Bayne, M. Cloning, expression and characterization of human alpha adrenergic receptors alpha-1A, alpha-1B, and alpha-1C. Biochem. Biophys. Res. Commun. 201: 1296-1304, 1994.
PubMed ID : 8024574 10 Chang, D. J.; Chang, T. K.; Yamanishi, S. S.; Salazar, F. H. R.; Kosaka, A. H.; Khare, R.; Bhakta, S.; Jasper, J. R.; Shieh, I.-S.; Lesnick, J. D.; Ford, A. P. D. W.; Daniels, D. V.; Eglen, R. M.; Clarke, D. E.; Bach, C.; Chan, H. W. Molecular cloning, genomic characterization and expression of novel human alpha-1A-adrenoceptor isoforms. FEBS Lett. 422: 279-283, 1998.
PubMed ID : 9490024 11 Assari T, Cox S, Munday MR, Pearce B. Regulation of alpha(1)-adrenoceptor-linked phosphoinositide metabolism in cultured glia: involvement of protein phosphatases and kinases. Cell Signal 15 (4): 403-12, 2003.
PubMed ID : 12618215 12 Khorchid A, Cui Q, Molina-Holgado E, Almazan G. Developmental regulation of alpha 1A-adrenoceptor function in rat brain oligodendrocyte cultures. Neuropharmacology 42 (5): 685-96, 2002.
PubMed ID : 11985827 13 Del Rio-Hortega, P. Estudios sobre la neuroglia. La glía de escasas radiaciones(oligodendroglía). Bo R Soc Espan Hist Nat; 21-63-92, 1921.
14 Cammermeyer, J. Morphologic distintions betwwen oligodendrocytes and microglia cells in the rabit cerebral cortex. Am J Anat; 118:227, 1966.
15 Du Y, Dreyfys F.C. Oligodendrocytes as provider of growth factors. J Neurosc Res 68:647-654, 2002.
PubMed ID:12111826 16 Clark RB, Perkins JB. Regulation of adenosine 3`,5`-cyclic monophosphate concentration in cultured human astrocytoma cells by catecholamines and histamine. Proc Nat Acad Sci USA 68:2757-2760, 1971.
PubMed ID:4330940 17 Gilamn AG, Nirenberg ;. Effect of catecholamines on the adenosine 3',5'-cyclin monophosphate concentrations of clonal satellite cells of neurons. Proc Natl Acad Sci USA 68:2165-2168, 1971.
PubMed ID:4332686 18 Stone EA, Ariano MA. Are glial cells targets of the central noradrenergic system? A review of the evidence. Brain Res Brain Res Rev 14 (4): 297-309, 1989.
PubMed ID : 2560410 19 Agullo L, Garcia A. Characterization of noradrenaline-stimulated cyclic GMP formation in brain astrocytes in culture. Biochem J 288 ( Pt 2): 619-24, 1992.
PubMed ID : 1334410 20 Kulik A, Haentzsch A, Luckermann M, Reichelt W, Ballanyi K. Neuron-glia signaling via alpha(1) adrenoceptor-mediated Ca(2+) release in Bergmann glial cells in situ. J Neurosci 19 (19): 8401-8, 1999.
PubMed ID : 10493741 21 Nalepa I, Kreiner G, Kowalska M, Sanak M, Zelek-Molik A, Vetulani J. Repeated imipramine and electroconvulsive shock increase alpha 1A-adrenoceptor mRNA level in rat prefrontal cortex. Eur J Pharmacol 2002 May 31;444(3):151-9.
PubMed ID : 12063075 22 Barrer BA, Raff MC. Proliferation of oligodendrocyte precursor cells depends on electrical activity in axons. Nature 361, 258-260, 1993.
PubMed ID:8093806 23 Demerens C., Stankoff B, Logak M, Anglade P, Allinquant B, Couranud F, Zale B, Lubertzki C. Induction of myelination in the central nervous system by electrical activity, Proc Nat Acad Sci USA 93:9887-9872, 1996.
PubMed ID:8790426 24 Khorchid A, Larocca JN, Almazan G. Characterization of the signal transduction pathways mediating noradrenaline-stimulated MAPK activation of c-fos expression in oligodendrocyte progenitors. J Neurosci Res 58; 765-778, 1999.
PubMed ID:10583908









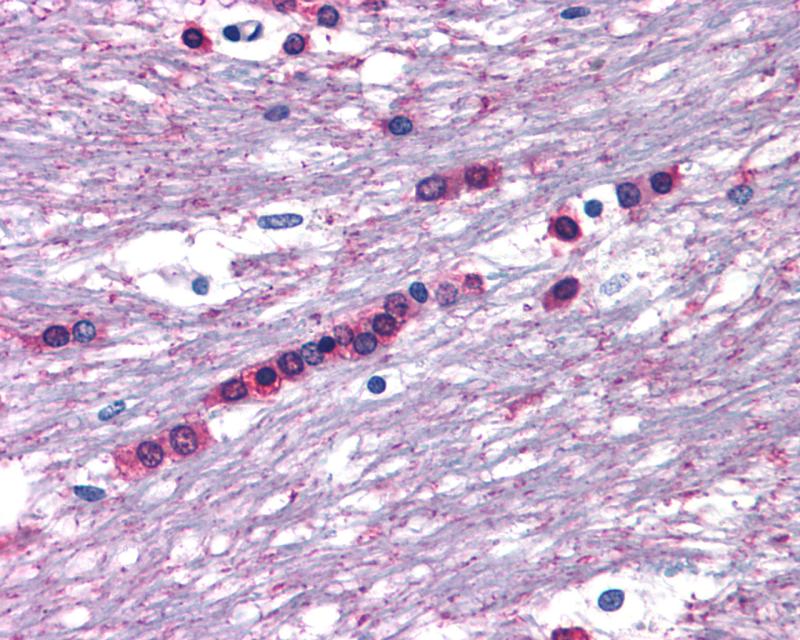 Zoom
Zoom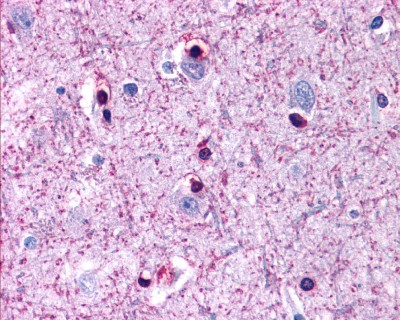
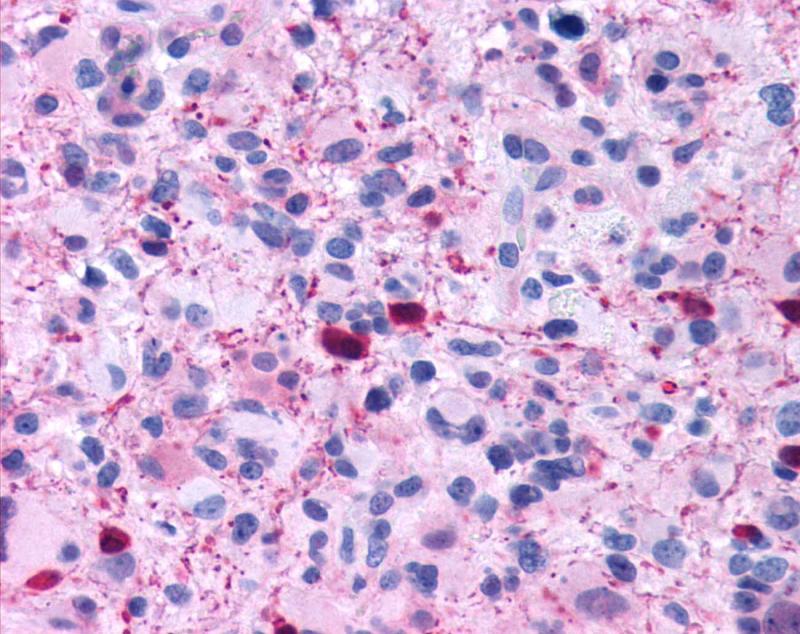 Zoom
Zoom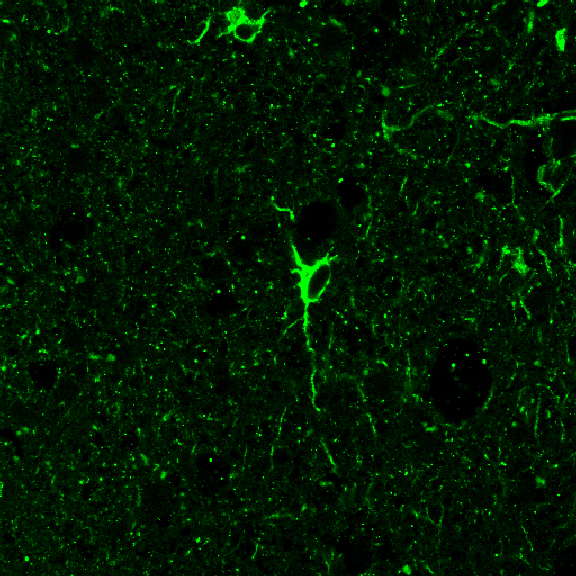 Zoom
Zoom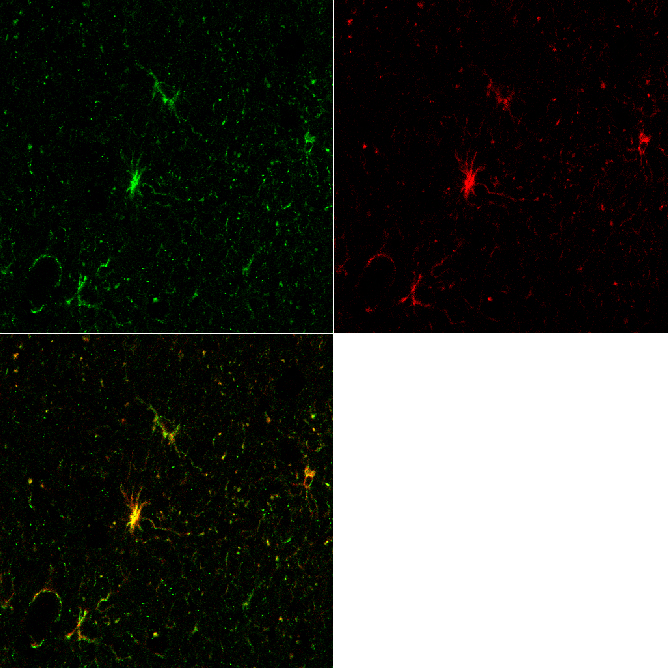 Zoom
Zoom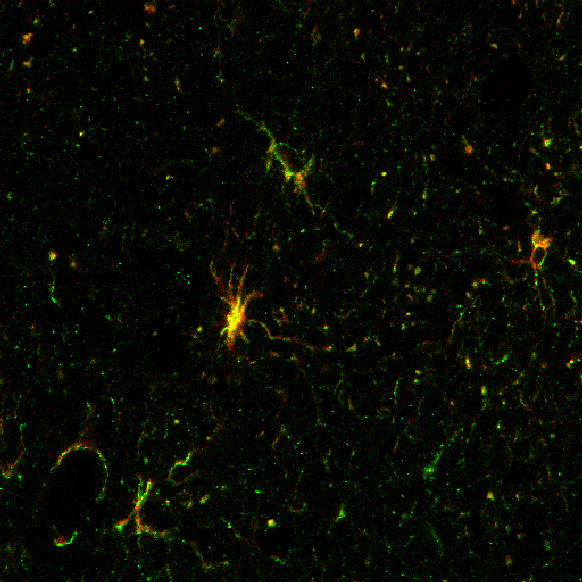 Zoom
Zoom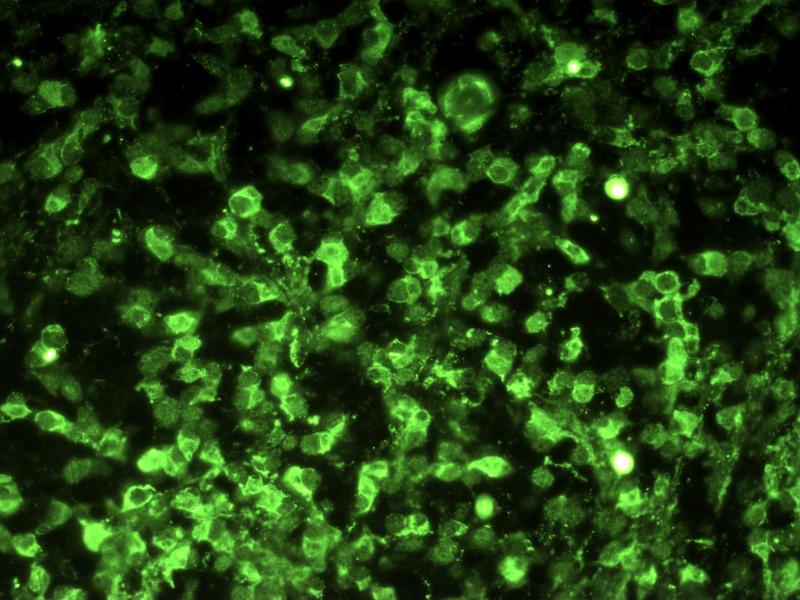 Zoom
Zoom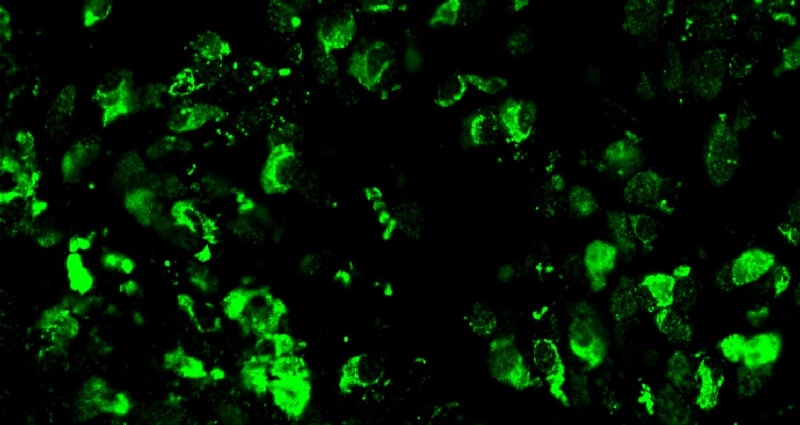 Zoom
Zoom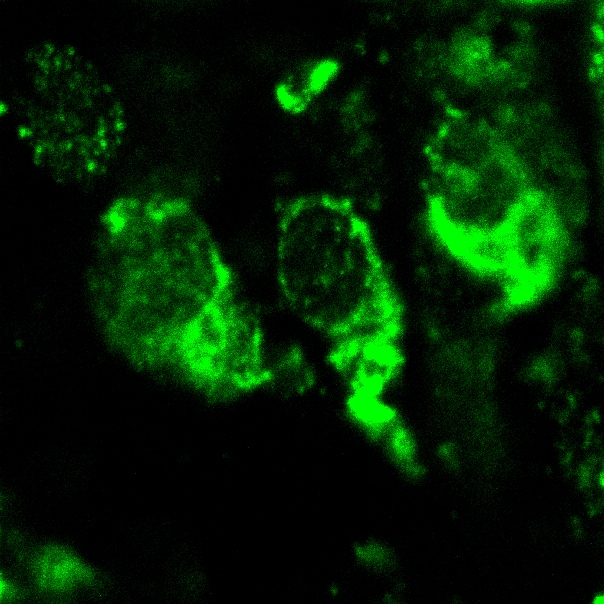 Zoom
Zoom