
ISBN: 978-84-692-76778
 COMUNICACIONES
COMUNICACIONES
Nº 2092. Patología Quirúrgica
|
Addys González Palomo[1], Armando López[2], Mariano Gabri[3], Daniel Alonso[4], Rolando Pérez[1], Enrique Montero[1] |
|
amlodipin krkaamlodipin 5 mg onlineA high EGFR expression was shown in the majority of tumor cells. An NGcGM3 ganglioside was immunodetected over the surface of the 3LL-D122 tumor cells. We found that both molecules widely co-localize on the lung metastases. Expression of EGFR and NGcGM3 were quantified in untreated mice by Intensity score. Moreover, we explored the hypothesis that the anti-metastatic effect of an anti-EGFR MAb may be modulate by the co-administration with an anti-NGcGM3 MAb, in vivo. Treatment of mice bearing 3LL-D122 with anti-EGFR MAb reduced the lung weight by decreasing the number of macrometastases and size of the micro-metastases on the pulmonary parenchyma. Anti-EGFR MAb decreased the cell proliferation and increased the apoptotic bodies. Anti-NGcGM3 MAb treatment decreased the mitotic and angiogenesis process without inducing apoptosis. Surprisely, treatment with anti-EGFR MAb plus anti-NGcGM3 MAb was more effective inhibiting pulmonary nodules than single-agent therapies. Histological examination revealed that combinatory therapy reduced the number of tumor foci and their size by decreasing the cell proliferation, increasing the apoptotic cells and this therapy inhibited the angiogenic process. Combinatorial targeted therapy to the EGFR and NGcGM3 ganglioside in C57BL/6 mice bearing the Lewis Lung Carcinoma increased the overall survival. These results suggest the anti-NGcGM3 ganglioside treatments may increase the anti-tumoral effect of the anti-EGFR therapy based in the co-expression of both molecules.
|
||
|
|
Non-small cell lung cancer (NSCLC) is the most common type of lung cancer and the development of metastasis is a major cause of death.
The EGFR-overexpression is observed frequently in NSCLC. EGFR has been intensively studied because it is a critical role in the growth and tumor cell proliferation. Lung cancer cells have diverse response to anti-EGFR therapies. Elucidation of cellular differential sensitivity holds the promise of better understanding the tumor growth biology.
In the last decades, the interactions of the EGFR with different gangliosides on the cell membrane have been described. However, the gangliosides are known to participate in the regulation of cell signaling within lipid rafts and/or caveolae by association with signaling proteins, such as Src family kinases . Furthermore, gangliosides have also been shown to regulate the activity of EGFR, fibroblast growth factor receptor. Different from other gangliosides, GM3 particularly may attenuate the EGFR signaling, inhibiting or increasing the EGFR dependent cell proliferation in a variety of cell types.
Depletion of endogenous GM3 inhibits the association of the uPAR/α5β1 integrin complex with EGFR but interestingly the uPAR (Urokinase-type plasminogen activator receptor), tends to be localized to lipid rafts. Recently, the GM3 synthase, SAT-1mRNA showed different expression levels in lung biopsy samples from NSCLC patients, contributing to the pathological identification of these kind of tumors. Additionally they found an increase in the EGFR protein levels and a gefitinib sensitivity in GM3-reconstituted cell suggesting the involvement of GM3 in the turnover of EGFR protein.
Therefore, the significance of the biological function of gangliosides in malignances is still controversial.
The role of the glycolilated NGcGM3 has been less studied in the tumor biology. NGcGM3 is expressed in human breast and melanoma tumors. On the other side, the immune response in individual immunized with a cancer vaccine containing an anti-idiotype mAb to gangliosides N-glycolilate have shown a substantial antitumor response.
Nevertheless, the association between expressions of NGcGM3 ganglioside with EGFR on tumor cells has not been studied yet. Moreover, there is a pressing need to unveil the mechanisms limiting the efficacy of anti-EGFR treatment in lung cancer patients.
In the present study, we examined the expression of EGFRand NGcGM3 ganglioside in lung metastases and the same time, we explored whether the anti-metastatic effect of anti-EGFR therapy would be modulated by the co-administration with an anti-NGcGM3 mAb, in vivo.
|
|
|
|
Animals: Eight-week-old female C57BL/6 mice (18-20 g) were purchased from the Center for Laboratory Animal Production (CENPALAB, Havana, Cuba). Animals were housed under conventional conditions with water and food ad libitum, maintained in accordance with the guidelines stipulated by Animal Subject Committee Review Board of the Centre of Molecular Immunology (CIM), Havana, Cuba.
Experimental metastases models: A single cell suspension of 2 x 105 of Lewis lung carcinoma (3LL-D122) in 0.5 mL of PBS was inoculated into a tail vein (designated as day 0 of the experiment). The mice were sacrificed on day 21 and pulmonary tissue was fixed by inflation through an intratracheal instillation of formalin. Then, the lungs were removed and fixed in Bouin´s fixative. The number of tumour colonies on the surface of each lung was counted under a dissecting microscope.
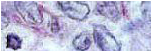
Histological analysis: Lungs were fixed in 10% neutral buffered formalin and it was processed by standard techniques. The tissue sections were stained with hematoxylin and eosin. Tumor cell proliferation (mitotic index) was assessed by the number of cells containing visible chromosomes divided by the total number of cells in the field of view. Apoptosis was quantified using the apoptotic index, similar to mitotic index. Ten random fields were evaluated in each tumor section from all animals per group.
Immunocytochemistry: The tumoral cells from animals without any treatment were removed from lung nodules using a glass homogenizer (Fisher Scientific, Pittsburgh, PA, USA) for immunocytochemistry assay (ICA) method. The 7A7 mAb was used to identify the EGFR (20). The 14F7 mAb was used to recognize specifically the NGcGM3 gangliosides (14). The primary antibody staining was detected by using a biotinylated rabbit anti-mouse immunoglobulin antiserum (DAKO, CA; USA). The immunoreaction was performed with the avidin-peroxidase complex (DAKO) and 3, 3-diaminobenzidine (DAB) (DAKO) as the chromogen. Sections were counterstained with hematoxylin. All observation was made by a light microscope (Olympus Optical CO., LTD, Tokyo; Japan).
Scoring system for immunocytochemistry: We evaluated the intensity (weak, moderate, or strong) and pattern (complete or incomplete) of membrane labelling in 10 high-power fields (X 400) of immunostaining. Combining intensity and membrane pattern of cells, score (0 to 3) was thus reached for each case. Tumor cells were considered negative (score 0) if cell membrane staining was completely absent, whereas a faintly appreciable and incomplete pattern (Score 1), a weak-to-moderate but complete staining (score 2), or a strong and complete labelling of the membrane in more than 10% in tumor cell were give (score 3). The expression index (total score) was obtained as follow: Intensity score ═ (% Score 1) + (% Score 2)x2 + (% Score 3)x3.
A known-positive control cell (A431 and X63 cells comparable in terms of EGFR and NGcGM3 expression, respectively) was included in each run of staining and consistently showed the expected immunoreactivity. Ten random fields were evaluated from each animals bearing 3LL-D122.
For endothelial cell detection by CD31/PECAM, all samples were incubated with 3% hydrogen peroxide in methanol (v/v) to block endogenous peroxidase, washed in PBS. Tissues sections were treated with 1% (w/v) normal goat serum. The sections were incubated with anti-CD31 MAb (Pharmigen, San Diego, CA, USA). Binding of the primary antibody was detected using biotinylated rabbit anti-mouse immunoglobulin secondary antiserum (Jackson Laboratory Bar Harbor ME, USA), followed by the avidin-biotin-peroxidase system (Dako, Denmark) with 3,3-Diaminobencidine (DAB) (Sigma, St. Louis, MO, USA) as the cromogen. Sections were counterstained with hematoxylin. Negative control involved the same procedure with the omission of the primary antibody. All observation was made by a light microscope (Olympus Optical CO., LTD, Tokyo; Japan).
Perimeters of endothelial cells: Tumor vessels-positive cells were evaluated by light microscopy, counted in five random fields containing exclusively viable tumoral cells, imaged digitally, and processed with ImageJ software (Version 1.24o).
Immunofluorescence study: Lung metastases were removed from animals non-treated. EGFR was immunolabeled with the byotinilated 7A7 mAb followed by streptavidin conjugated to FITC. NGcGM3 ganglioside was identified with the 14F7 mAb and rhodamine conjugated byotinilated anti-mouse IgG. The immunostaining pattern was analyzed with a confocal laser microscope Fluoview FV500 TM (Olympus, Tokyo; Japan).
Statistical analyses: The relative weight, number of metastatic tumor colonies and mitotic/apoptotic tumor index were compared by Kruskall Wallis test. Ten random fields were evaluated from each animals bearing 3LL-D122. Values were considered statistically significant when p<0.05.
|
|
|
|
A high EGFR expression was shown in the majority of tumor cells. An NGcGM3 ganglioside was immunodetected over the surface of the 3LL-D122 tumor cells. We found that both molecules widely co-localize on the lung metastases. Expression of EGFR and NGcGM3 were quantified in untreated mice by Intensity score (Fig 1). Treatment of mice bearing 3LL-D122 with anti-EGFR MAb reduced the lung weight by decreasing the number of macrometastases and size of the micro-metastases on the pulmonary parenchyma. Anti-EGFR MAb decreased the cell proliferation and increased the apoptotic bodies. Anti-NGcGM3 MAb treatment decreased the mitotic and angiogenesis process without inducing apoptosis. Surprisely, treatment with anti-EGFR MAb plus anti-NGcGM3 MAb was more effective inhibiting pulmonary nodules than single-agent therapies. Histological examination revealed that combinatory therapy reduced the number of tumor foci and their size by decreasing the cell proliferation, increasing the apoptotic cells and this therapy inhibited the angiogenic process (Fig 2). Additionally, overall survival in animals treated with the Combination therapy was significantly higher (Fig 3).
|
|
|
|
The significance of the relation between GM3 and EGFR is still controversial, although there are some reports which it has demonstrated the co-localization of EGFR in the lipid rafts and/or caveolae (Parton 1994, A. 2001) with different gangliosides (Mobius W. 1999). Caveola have proposed to be the site of EGFR signaling including EGFR autophosphorylation, based-on the demonstration that caveolin-1 and EGFR both co-localize on the membrane. (Pike LJ., Miller JM 1998).
In the present study, we showed EGFR expression on lung metastasis induced by 3LL-D122 cell line and it is in accordance with others reports which showed that EGFR are highly expressed in many tumors of epithelial origin (Mayumi Ono, 2004; Bradender J, 2001). A similar result has been reported for the studies in H125, non-small cell lung carcinoma, which showed a high EGFR levels (Christensen JG., et al. 2005).
Additionally, in this study, the NGcGM3 ganglioside was detected in tumor cell removed from lung metastasis. In contrast, the staining pattern was different to EGFR expression. Cell membranes that were intensely stained to NGcGM3 were visible and distributed non homogeneous pattern over the surface of the 3LL-D122 cells.
Surprisingly, we found that NGcGM3 ganglioside co-localized with EFGR in the same tumor cells.
Our result evidence the presence of NGcGM3 in lung metastasis. This fact has been reported in other types of tumor for example: Melanoma and breast cancer (Oliva JP, 2006). However, EGFR and NGcGM3 co-expressed on the 3LL-D122 tumor cell and this may to offer the opportunity, in vivo, to investigate the functions of endogenous gangliosides in tumor biology.
Some studies have shown tumor cell resistance to strategies with multiple anti-EGFR therapies. These resistance mechanisms are mediated through other glycoprotein that initiate parallel signal transduction pathways and promote cell growth and proliferation despite the EGFR blockade (Contessa JN., 2008).
In this work, the co-localization of the EGFR with the NGcGM3 from experimental metastasis model induced by the Lewis lung carcinoma, provides the opportunity to evaluate in an animal models, NGcGM3 in lung cancer and if the anti-ganglioside therapy could enhance the sensitivity of this tumor to anti-EGFR treatments.
Here, we explored the possibility that antibody against NGcGM3 ganglioside (14F7 mAb) might modulate the anti-EGFR therapy (7A7 mAb), to improved its anti-tumor effects. Using the experimental lung metastasis model, we found that the combination therapy was superior to the individual drugs on the metastasis induced by 3LL-D122 cells. Our observations indicated the reduction of the relative pulmonary weight and significantly decreased the number of macrometastasis and micrometastasis in the pulmonary parenchyma. Additionally, the nodules from animals treated with the combinatory therapy showed the smaller size of micro nodules.
These data collectively suggest that although anti-EGFR showed the anti-metastatic effect as it was reported (Greta 2004), their result was increased by the co-administration of the anti-ganglioside therapy as we showed here.
To conclude, our findings suggest that co-expression of NGcGM3 ganglioside with the EGFR on tumor tissues may be useful to continue studying the functional role of GM3 ganglioside signaling domains.
Several mechanisms of ganglioside action have been proposed (Li R.,2000, Contessa JN., 2008). The challenge now is to define exactly if the effect of anti-NGcGM3 mAb is direct or indirect on 3LL-D122 cell. In case of direct effect will be to evidence the oncosis phenomenon (Roque-Navarro, 2007) or to demonstrate the ability of anti-NGcGM3 mAb may regulate the movement of the extracellular domain of EGFR in the lipid raft allowing the functional signaling at cellular membrane (Li R.,2000).
Indirect effect will be address by the castration of ganglioside by the anti-NGcGM3 mAb in the tumor microenvironment allowing the movement of the lipid rafts. Additionally, the capacity of ganglioside to down-modulate to CD4 expression in murine and human T-lymphocytes on this tumor was analyed (De Leon J., 2007). Additionally, overall survival in animals treated with the Combination therapy was significantly higher
|
|
|
|
In summary te combination of targeting the EGFR and NGcGM3 was effective in significantly inhibiting tumour growth and the overall survival in this experimental model. Targeting multiples pathways strategy may be a potential treatment strategy for lung cancer that are dependent on EGFR and NGcGM3 for tumor proliferation, angiogenesis and survival.
|
|
|
|
1- Kraus-Berthier L, Jan M, Guilbaud N, Naze M, Pierré A, Arassi G. Histology and sensitivity to anticancer drugs lung carcinomas implanted in the pleural cavity of nude mice. Clin Cancer Res. 2000; 6: 297-304.
2- Suzuki M, Sari ME, Montel V, Tarin D. Dormant cancer cells retrieved from metastasis-free organs regain tumorigenic and metastatic potency. Am Journal Pathol. 2006; 169: 2: 673-81.
3- Bruns CJ, Harbison MT, Davis DW, Portera CA, Tsan R, McConkey DJ, et al. Epidermal growth factor receptor blockade with C225 plus gemcitabine results in regression of human pancreatic carcinoma growing orthotopically in nude mice by antiangiogenic mechanisms. 2000; 6: 1936-48.
4- Ono M, Huwano M. Molecular mechanisms of epidermal growth factor receptor (EGFR) activation and response to gefitinib and other EGFR-targeting drugs. Clin Cancer Res 2006; 12: 7242-51.
5- Noguchi M, Kabayama K, Uemura S, Kang Bw, Saito M, Igarashi Y. et al. Endogenously produced ganglioside GM3 endows etoposide and doxorubicin resistance by up-regulating Bcl-2 expression in 3LL Lewis lung carcinoma cells. Glycobiology. 2006; 16: 641-50.
6- Zhang Q, Furukawa K, Chen H, Sakakibara T, Urano T, Furukawa K. Metastatic Potential of mouse Lewis Lung Cancer Cells is regulated via ganglioside GM1 by modulating the matrix metalloprotease-9 localization in lipid rafts. J. Biol. Chem. 2006; 281:26: 18145-55.
7- De León Y, Fernández A, Mesa C, Clavel M, Fernández LE. Role of tumor-associated N-glycolylated variant of GM3 ganglioside in cancer progression: effect over CD4 expression on T cells. Cancer Immunol. Immunothera. 2006, 55: 443-50.
8- Livingston PO, Zhang S, Lloyd KO. Carbohydrate vaccines that induce antibodies against cancer: 1. Rationale. Cancer Immunol. Immunother. 1997; 45: 1-9, (suppl 1).
9- Furuka K, Yamaguchi H, Oettgen, HF, Old LJ, Lloyd KO. Analysis of the expression of N-Glycolylneuraminic Acid-containing gangliósidos in cell and Tissues using two human monoclonal antibodies. J. Biol. Chem. 1988; 263: 18507-12.
10- Nohara K, Wang F, Spiegel S. Glycosphingolipid composition of MDA-MB-231 and MCF-7 human breast cancer cell lines. Breast Cancer Res. Treat. 1998; 48: 149-57.
11- Deng W, Li R, Ladisch S. Influence of cellular ganglioside depletion on tumor formation. J. Natl. Cancer Inst. 2000; 92: 912-17.
12- Wang XQ, Sun P, Paller AS. Ganglioside GM3 blocks the activation of epidermal growth factor receptor induced by integrin at specific tyrosine sites. J.Biol.Chem. 2003; 278: 48770-8.
13- Oliva JP, Valdés Z, Casacó A, Pimentel G, González J, Álvarez I, Osorio M. et al. Clinical Evidence of GM3 (NeuGc) ganglioside expression in human breast cáncer using the 14F7 monoclonal antibody labelled with 99mTc. Breast Cancer Res. and Treatment. 2006, 96: 115-121.
14- Carr A, Mullet A, Mazorra Z, Vázquez AM, Alfonso M, Mesa C. et al. Mouse IgG1 monoclonal antibody specific for N-glycolyl GM3 ganglioside recognized breast and melanoma tumors. Hybridoma. 2000; 19: 241-47.
15- Miljan EA, Meuillet EJ, Mania-Farnell B et.al. Interaction of the extracellular domain of the epidermal growth factor receptor with gangliosides. J. Biol. Chem. 2002.
16- Wang XQ, Sun P, Paller AS. Ganglioside induces caveolin-1 redistribution and interaction with the epidermal growth factor receptor. J. Biol. Chem. 2002; 277: 47028-34.
17- Song W, Vacca MF, Welti R, Rintoul DA. Effects of ganglioside GM3 and De-N acetyl GM3 on Epidermal Growth Factor Receptor kinase Activity and Cell growth. J. Biol. Chem. 1991; 266: 5: 10174-181.
18- Sottocornola E, Berra B, Colombo I. GM3 content modulates the EGF-activated p 185c-neu levels, but not those of the constitutively activated oncoprotein 185c-neu. Biochim. Biophys. Acta. 2003; 1635:55-60.
19- Garrido G., Sánchez B, Rodríguez HM, Lorenzano P, Alonso D, Fernández LE. 7A7 MAb. A new tool for the pre-clinical evaluation of EGFR-based therapies. Hybridoma and Hybridomics. 2004; 23: 168-75.
20- Okabe T, Okamoto I, Tamura K. Differential constitutive activation of the epidermal growth factor receptor in non-small cell lung cancer cells bearing EGFR gene mutation and amplification. Cancer Res. 2007; 67: 2046-53.
21- Ihde D, Chemotherapiy lung cancer. New England J.Med, 1992; 327:1434-41
22- De Pas Tommaso, Pelosi G, de Braud F, Veronesi G, Curigliano G, Leon ME. Et al. Modulation of epidermal growth factor receptor status by chemotherapy in patients with locally advanced non-small-cell lung cancer is rare. J Clin Oncol. 2004; 22: 4966-70.
23- McLemore T, Eggleston J, Shoemaker R, Abbot B:J:, Bohlman M, Liu M, Fine D:L., Mayo J:G:, Boyd M:R: Comparison of intrapulmonary, percutaneous intrathoracic and subcutaneous models for the propagation of human pulmonary and nonpulmonary cancer cells lines in athymic nude mice. Cancer Res., 48: 2880-2886, 1988.
24- Manzotti C., Audisio R., Patresi G. Importance of ortho-topic implantation for human tumors as model systems: relevance to metástasis and invasión.Clin. Exp. Metastasis, 11: 5-11, 1993.
25- Nakanishi H, Takenaga K, Oguri K, Yoshida A, Okayama M. Morphological characteristics of tumours formed by Lewis lung carcinoma-derived cloned cell lines with different metastatic potentials: structural differences in their basement membranes formed in vivo. Virchows Archiv A Pathol Anat. 1992; 420: 163-70
26- Bagheri-Yarmand R, Kourbali Y, Rath AM et al. Carboxymethyl benzylamide dextran blocks angiogenesis of MDA-MB435 breast carcinoma xenografted in fat pad and its lung metastases in nude mice. Cancer Res. 1999; 59: 507-10.
27- Fernández LE, Alonso DF, Gómez DE, Vázquez AM. Ganglioside-based vaccines and anti-idiotype for active immnunotherapy against cancer. Expert Rev. Vaccines. 2003; 2: 89-95.
28- Carr A, Mesa C, Arango MC, Vázquez AM. et al. In Vivo and In Vitro anti-tumor effect of 14F7 monoclonal antibody. Hybridoma and Hybridomics, 2002; 21: 463-68
29- Watanabe R, Ohyama C, Aoki H, Takahashi T, Satoh M, Saito S. et al. Ganglioside GM3 overexpression indices apoptosis and reduces malignant potential in murine bladder cancer. Cancer Res. 2002; 62: 3850-54.
30- Carr A, Rodríguez E, Arango MC, Camacho R. Immunotherapy of Advance Breast cancer with a heterophilic ganglioside (NeuGcGM3) cancer vaccine. J Clin Oncol. 2003; 2: 1015-21
31- Díaz A, Alfonso M, Alonso R, Suarez G, Troche M, Catalá M. et al. Immune response in breast cancer patients immunized with an anti-idiotype antibody mimicking neu-Gc-containing ganglioside. Clinical Immunol. 2003; 107: 80-89.
32- Díaz A, Alfonso M, Alonso R, Suarez G, Troche M, Catalá M. et al. Immune response in breast cancer patients immunized with an anti-idiotype antibody mimicking neu-Gc-containing ganglioside. Clinical Immunol. 2003; 107: 80-89.
33- Epidermal growth factor receptor mutations and susceptibility to targeted therapy in lung cancer. Respirology. 2006; 11: 687-92.
34- Ishii A, Ohta M, Watanabe Y, Matsuda K, Ishiyama K, Sakoe K, et. al. Expression cloning and functional characterizarion of human cDNA for gangliosides GM3 synthase. J. Biol. Chem. 1998; 273: 31652-55.
35- Noguchi M, Suzuki T, Kabayama K, Takahashi H, Chiba H, Shiratori M. et al. GM3 synthase gene is a novel biomarker for histological classification and drug sensitivity against epidermal growth factor receptor tyrosine kinase inhibitors in non-small cell lung cancer. Cancer Sci. 2007; 98: 1625-32.
36- Wang XQ, Sun P, Paller AS. Gangliosides inhibit urokinase-type plasminogen activator (uPA)-dependent squamous carcinoma cell migration by preventing uPA receptor/α5β1 Integrin/Epidermal Growth Factor Receptor interactions. J. Invest. Dermatol. 2005; 124: 839-48.
37- Nicholson RI, Gee JM, and Harper ME. EGFR and cancer prognosis. Eur J Cancer 2001; 37:S9-15
38- Irie RF, Irie K, Morton DL. A membrane antigen common to human cancer and fetal brain tissues. Cancer Res.36, 3510-3517 1976).
39- Li R, Manela J, Kong Y and Ladisch S, Cellular gangliosides promote growth factor-induced proliferation of fibroblasts. 2000, J Biol Chem 275, 34213-34223.
40- Mayumi Ono, Akira Hirata, Takuro Kometani, Miho Miyagawa, Shu-ichi Ueda, Hisafumi Kinoshita, Teruhiko Fuji and Michihiko Kuwano, Sensitivity to gefitinib (Iressa, ZD1839) in non-samll cell lung cancer cell lines correlates with dependence on the epidermal growth factor (EGF) receptor/extracellular signal-regulated kinase ½ and EGF receptor/AkT pathways for proliferation. Molecular Cancer Therapeutics 2004; 3 (4): 465-472.
41- Danan Li, Hongbin Ji, Sara Zaghlul, Kate McNamara, Mei-Chic Liang, takeshi Shimamura, Shigeto Kubo, Masaya Takahaschi, Lucian R. Chirieac, Robert F. Padera, Andrew M. Scot, Achim A. Jungbluth, Webster K. Cavenee, Lloyd J. Old, George D. Demetri and Kwok-Kin wong. 2007, J. Clin.Invest. 117:346-352.
42- Hofman EG, Ruonala MO, Bader AN, van den Heuvel D, Voortman J, Roovers RC. EFD induces coalescence of different lipid rafts. J Cell Sci. 2008.Aug 1; 121 (15): 2519-28.
43- Loo D, Pryer N, Young P, Liang T, Coberly S, King KL, Kang K, Roberts P, Tsao M, Xu X, Potts B, Mather J. The glycotope-specific RAV12 monoclonal antibody induces oncosis in vitro and has antitumor activity against gastrointestinal adenocarcinoma tumor xenografts in vivo. Mol Cancer Ther 2007; 6(3) March 856-865.
44- Christensen JG, Vincent PW, Klohs WD, Fry DW, Leopold WR, Elliott WL. Plasma vascular endothelial growth factor and interleukin-8 as biomarkers of antitumor efficacy of a prototypical erbB family tyrosine kinase inhibitor. Molecular Cancer Therapeutics. 2005; 4(6) June, 938-947.
45- Xu Y, L.J.Tan, Grachtchouk, J.J.Voorhees, G.J.Fisher. Receptor-type protein-tyrosine phosphate-kappa regulates epidermal growth factor receptor function. 2005. J.Biol. Chem. 280, 42694-42700
46- Jong-Shiaw J., Dar-Shih H., Shih-Hurng L., Chen A., Chen-Wen Y and Chung-Yang Y. Increasing expression of serine protease matriptase in ovarian tumors: tissue microarray analysis of immunostaining score with clinicopathological parameters. Modern Pathology. 2006; 19: 447-452
47- S-M Kin, Ji-Ung J, Jae-Sung R, Jung-Woo J, Hyo-Jung Y, Kisung Ko, Hyung-Keun Y, Kyu-Yong J, Young-Kug c: Effect of gangliosides on the differentiation of human mesenchymal stem cell into osteoblasts by modulating epidermal growth factor receptors. Biochemical and Biophysical Research Communications 2008; 307: 806-871.
48- Contessa JN., Bhojani MS., Freeze HH., Rehemtulla A., Lawrence TS. Inhibition of N-linked glycosilation disrupts receptor tyrosine kinase signaling in tumor cells. Cancer Res. 2008; 68 (10): May 15; 3803-3809.
|
|
|
|
- Isidro Machado - ESPAÑA (05/11/2009 0:58:21)
- Daniela Ancich - ARGENTINA (06/11/2009 19:07:02)
- Addys Gonzalez Palomo - CUBA (Autor) (09/11/2009 17:28:49)
- Isidro Machado - ESPAÑA (11/11/2009 12:54:54)
|
|
|
|
|
.JPG)
.JPG)



Web mantenido y actualizado por el Servicio de informática uclm. Modificado: 16/06/2015 11:37:37